


Autophagie
Macroautophagie
Formation de l'autophagosome
Élongation du phagophore
: apport de lipides et courbure
- Vue d'ensemble du système endomembranaire
- Autophagie
- Système UPS (Système UPS (Ubiquitine-protéasome))
- Fusion et fission membranaire
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

L'élongation du phagophore se fait principalement par l'incorporation de lipides, et sa courbure membranaire est affectée par plusieurs facteurs.
L'élongation et la courbure membranaire impliquent :
- deux systèmes de conjugaison de type ubiquitine, le système ATG12 et le système ATG8,
- une expansion membranaire par l'apport de lipides.

Vous pouvez lire : Autophagosome biogenesis: From membrane growth to closure (2020), Emerging roles of ATG proteins and membrane lipids in autophagosome formation (2020) et Structural view on autophagosome formation (2023), ce dernier article analyse les structures des différents composants.
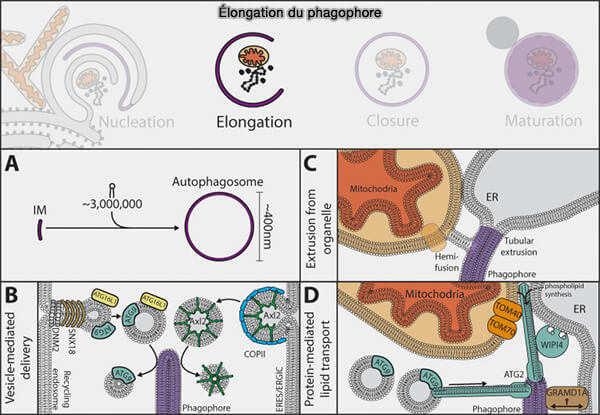
(Figure : vetopsy.fr d'après Melia et coll)
Expansion membranaires par les lipides
Vue d'ensemble
L’autophagie nécessite la production de grandes quantités de membranes biologiques à des échelles physiques relativement grandes et sur des délais courts.
1. Comme le phagophore en expansion est en grande partie dépourvu de protéines transmembranaires, l'expansion membranaire se fait principalement par l'incorporation de lipides (Autophagosome biogenesis: From membrane growth to closure 2020).
- En réponse au stress de la macroautophagie, la mobilisation pourrait atteindre plus de 100 000 000 lipides par cellule.
- Pour ce faire, le réticulum endoplasmique (RE) augmente la synthèse lipidique (
 ULK et PIS).
ULK et PIS).
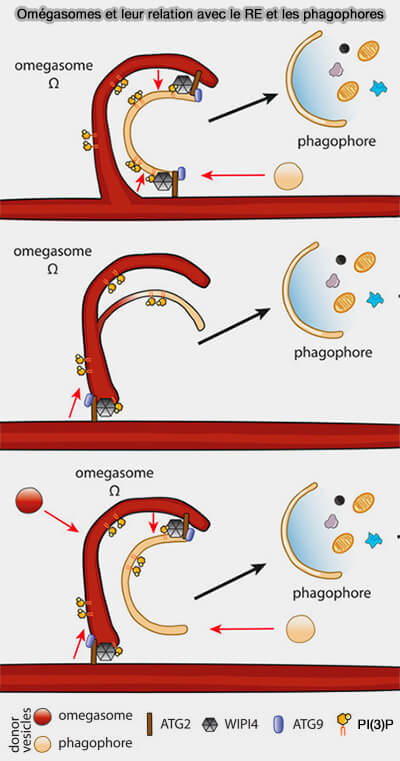
(Figure : vetopsy.fr d'après Norell et coll)
2. La distribution des lipides dans les omégasomes diffère selon les études, i.e. les omégasomes sont connectés ou pas avec le réticulum endoplasmique (RE).
a. Les études antérieures suggéraient que les omégasomes sont des domaines du RE qui agissent comme des plates-formes pour la formation de novo des autophagosomes.
Les lipides nécessaires à la nucléation et à l’expansion des phagophores sont fournis par les vésicules ou par transfert direct de lipides depuis le RE et/ou les omégasomes par ATG2.
b. Des études récentes ont suggéré que les omégasomes ne sont pas connectés au RE, ce qui indique qu’ils sont clivés du RE après leur formation.
Les phagophores peuvent émerger par un mécanisme de bourgeonnement par remodelage des membranes omégasomales, i.e. le RE peut donner des lipides par transfert direct médié par ATG2.
c. Les omégasomes peuvent également se former par fusion vésiculaire et par transfert de lipides à partir du RE.
3. Les lipides semblent atteindre le phagophore en expansion par trois processus :
- le transport direct par protéines des lipides,
- l'approvisionnement par des vésicules,
- l'extrusion directe à partir d'un organite préexistant.
Transport direct par des protéines
Au moins trois LTP (Lipid Tranfer Protein) ou protéines de transfert des lipides sont maintenant connues pour soutenir la biogenèse des autophagosomes.
Les LTP (Lipid Tranfer Protein) ou protéines de transfert des lipides ont la capacité de déplacer les lipides entre les membranes, dans lesquelles ils sont distribués de manière très asymétrique.
ATG2

Les différentes LTP, et en particulier ATG2 ou VPS13, sont étudiées dans des chapitres spéciaux.
1. Des protéines à domaine choréine telles que ATG2 ou VPS13, protéines à domaine choréine, peuvent relier deux membranes par un long pont protéique de 16 nm en forme de tunnel avec une rainure hydrophobe à travers laquelle les lipides peuvent glisser, alors que la protéine reste stationnaire.
ATG2 engage chacune de ces membranes (ATG2 transports lipids to promote autophagosome biogenesis 2019 et Atg2A/B deficiency switches cytoprotective autophagy to non-canonical caspase-8 activation and apoptosis 2017).
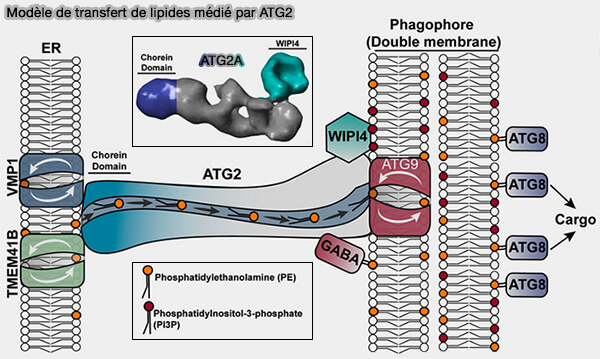
(Figure : vetopsy.fr d'après MacEwan et coll)
2. Le recrutement spécifique d'Atg2/ATG2B au phagophore nécessite des partenaires ( partenaires de liaison d'ATG2) :
à son extrémité N-terminale :
à son extrémité N-terminale :
- Atg9 par un hétérotétramère,
- WIPI4/Atg18, par le domaine CAD (Cys-Ala-Asp ou YFS) d'ATG2 sur les membranes enrichies en PI(3)P du phagophore ( interactions ATG2/WIPI4),
- GABARAP, par son domaine LIR.
Le recrutement d'ATG2B et de WIPI4 au phagophore dépend de l'extrémité C-terminale de TRAPPC11 du complexe TRAPP-III ( complexe TRAPP-III et autophagie).
3. En outre, ATG2A interagit avec TOM40/TOM70 sur les mitochondries ( interactions ATG2/TOM40).
Remarque : ATG2 et TRAP-III sont nécessaires également à la fermeture de l'autophagosome ( rôle d'ATG2 dans la fermeture de l'autophagosome et partenaires d'Atg2)
GRAMD1A
GRAMD1A semble être directement impliqué dans la biogenèse des autophagosomes par la régulation de l’homéostasie du cholestérol au niveau des phagophores/autophagosomes (The cholesterol transfer protein GRAMD1A regulates autophagosome biogenesis 2019).
GRAMD1A est une protéine de transfert du cholestérol de la famille du domaine StART dont on a découvert récemment le rôle dans l'autophagie (Toward the role of cholesterol and cholesterol transfer protein in autophagosome biogenesis 2019).
1. Lors de l’induction de l’autophagie, GRAMD1A s’accumule aux sites d’initiation des autophagosomes au réticulum endoplasmique (RE), potentiellement en raison de l’enrichissement en PI(3)P dans ces sites.
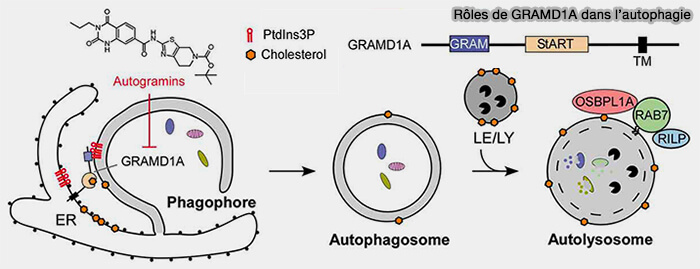
(Figure : vetopsy.fr d'après Wu et coll)
2. De même, la teneur en cholestérol des membranes autophagosomales augmente lors de la maturation de l'autophagosome, et en partie à cause de la fusion avec les endosomes tardifs/lysosomes riches en cholestérol via OSBPL1A/ORP1L.
Remarque : les autogramines sont des inhibiteurs de l'autophagie en concurrençant directement la liaison au cholestérol dans le domaine StART de GRAMD1A.
Approvisionnement par des vésicules : Atg9
Atg9/ATG9 est localisée dans des vésicules provenant de l'appareil de Golgi proches de l'autophagosome en développement.
L'Atg9 est étudié dans un chapitre spécial.
1. Chez la levure, ces vésicules peuvent s'agglutiner avec les complexes Atg1 pour conduire à l'initiation de la biogénèse des autophagosomes (The Atg1–kinase complex tethers Atg9-vesicles to initiate autophagy 2016).
Ce processus s'effectue dans une structure distincte par une séparation de phase liquide-liquide ou LLPS (Phase separation organizes the site of autophagosome formation, 2020).
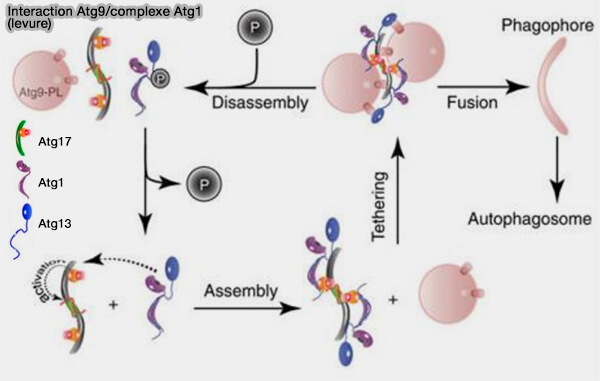
(Figure : vetopsy.fr d'après Rao et coll)
a. Les deux protéines Atg17 et Atg11 sont des composants d’échafaudage du complexe kinase Atg1.
- Atg17 recrute et attache les vésicules donneuses d’Atg9 dans la macrophagie non sélective ( interactions Atg17/Atg9).
- Atg11 initie l’autophagie sélective et coordonne le passage à l’autophagie non sélective en recrutant Atg17( Atg11 et autophagie sélective chez la levure).
b. Ensuite, la fusion homotypique de ces vésicules donne naissance au phagophore (Atg9 vesicles are an important membrane source during early steps of autophagosome formation 2012).
2. Chez les mammifères, le phagophore est entouré de tubules et de vésicules qui peuvent accepter ATG9, mais bien que les vésicules ATG9 soient essentielles à la phase d'expansion, il est probable que les lipides sont également récoltés à partir d'autres sources (Autophagosome biogenesis: From membrane growth to closure 2020).
- Le flux de vésicules ATG9 vers et depuis le phagophore pendant l'expansion de la membrane peut servir à des fins supplémentaires, telles que l'échafaudage de complexes protéiques fonctionnant au lancement de la biogénèse de l'autophagosome et l'organisation des machines nécessaires lors de l'expansion telle qu'ATG2 ( ATG2 et transfert lipidique vers le phagophore).
3. Cependant, une étude récente suggère qu'ATG9 se lierait aussi directement à LC3 dans les phagophores et aurait la capacité de promouvoir la formation des autophagosomes (Mammalian ATG9s drive the autophagosome formation by binding to LC3 2020).
Remarque : des vésicules COPII et Axl2 se colocalisent avec des phagophores en croissance, soutenant un rôle direct pour ces vésicules dans l'expansion du phagophore (COPII vesicles contribute to autophagosomal membranes 2019).
- Des sites de sortie uniques du RE pourraient produire des vésicules dédiées à la formation des autophagosomes ou aux processus liés à l'autophagie.
- En contrôlant la formation des vésicules destinées à l'expansion de l'autophagosome, ces sites RE modifiés pourraient également réguler l'accessibilité des protéines transmembranaires, fournissant une explication potentielle de la pénurie relative de ces molécules sur l'organite mature.
Extrusion directe à partir d'un organite préexistant
1. Les phagophores se forment à partir des omégasomes, sous-domaines du réticulum endoplasmique (RE).
a. La tomographie par cryo-électron et la microscopie électronique immuno-ordonnée suggèrent une éventuelle continuité entre le phagophore en forme de coupe et le RE environnant.
- La croissance pourrait alors se faire par extrusion continue et reposer sur l'apport quasi illimité de lipides par le RE.
- De plus, les composants précoces de la machinerie autophagique sont présents aux omégasomes, et plusieurs protéines localisées au RE ont été impliquées dans la biogenèse des autophagosomes (ER platforms mediating autophagosome generation 2020).
b. À l'inverse, des études n'ont pas pu observer les liens directs avec le RE, i.e. la généralité d'extrusion du RE reste incertaine.
2. La biogenèse de l'autophagosome a été documentée sur d'autres sites que le RE, au niveau :
- des mitochondries (Mitochondria supply membranes for autophagosome biogenesis during starvation 2011),
- des endosomes de recyclage (The RAB11A-Positive Compartment Is a Primary Platform for Autophagosome Assembly Mediated by WIPI2 Recognition of PI3P-RAB11A 2018).
Les omégasomes chevauchent les sites de contact RE/mitochondrie et les sites de sortie RE spécifiques, où les vésicules ATG9 semblent également être recrutées, suggérant une interconnexion étroite entre plusieurs sources de membrane de biogenèse des autophagosomes (Autophagy initiation by ULK complex assembly on ER tubulovesicular regions marked by ATG9 vesicles 2016).
Courbure membranaire
La modélisation théorique de la biogenèse des autophagosomes suggère que le phagophore se développe sous la forme d'une feuille à membrane aplatie qui, après la relaxation de l'énergie de la courbure de la membrane, se plie en un autophagosome sphérique (Formation of autophagosomes coincides with relaxation of membrane curvature 2019).
Cette étude suggère l'existence de mécanismes pour éviter une flexion prématurée du phagophore, plutôt qu'une machine pour conduire leur flexion.
Les courbures membranaires sont étudiées dans un chapitre spécial.
Plusieurs facteurs peuvent affecter la courbure membranaire.
1. PI(3)P et la phosphatidyléthanolamine (PE) sont des lipides en forme de cône ( courbures membranaires et changements lipidiques).
Le regroupement de PI(3)P au niveau du phagophore peut créer un bourgeon faisant face au cytosol dans la membrane qui sert de plate-forme pour le recrutement de la machinerie autophagique, via WIPI2 (WIPI2 Links LC3 Conjugation with PI3P, Autophagosome Formation, and Pathogen Clearance by Recruiting Atg12–5-16L1 2014).
2. La courbure membranaire des phagophores peut également être affectée par la liaison de diverses protéines.
a. On peut citer des protéines à domaine BAR comme :
- la nexine SNX18 (Membrane remodeling by the PX-BAR protein SNX18 promotes autophagosome formation 2013),
- BIF-1/Endophiline B1 (Bif-1 regulates Atg9 trafficking by mediating the fission of Golgi membranes during autophagy 2011),
- les protéines de fission Atg20 et Atg24 de la levure (Atg20- and Atg24-family proteins promote organelle autophagy in fission yeast 2016).
b. Plusieurs composants des complexes ULK et PIK3C3 contiennent des régions spécifiques qui facilitent probablement le recrutement membranaire de manière géométrique :
- un domaine EAT dans Atg1/ULK1 (The dynamic Atg13-free conformation of the Atg1 EAT domain is required for phagophore expansion 2018),
- un domaine BATS dans ATG14.
c. La conjugaison covalente à un lipide, i.e. lipidation de ATG8/LC3/GABARAP à la phosphatidyléthanolamine (PE) médiée par ATG12-ATG5-ATG16L1, est également très sensible à la courbure (Distinct functions of ATG16L1 isoforms in membrane binding and LC3B lipidation in autophagy-related processes 2019).
Elle est plus efficace sur les liposomes à haute courbure (25-65 nm) que sur ceux dont la courbure est relativement faible (plus de 400 nm), probablement dû aux hélices amphipathiques (AH) de :
- l'E2-enzyme ATG3 qui facilite la lipidation préférentiellement sur les membranes avec des défauts locaux de l'emballage lipidique (Lipidation of the LC3/GABARAP family of autophagy proteins relies upon a membrane curvature-sensing domain in Atg3 2014),
- ATG16L1 est également requise pour sa liaison membranaire et sa fonction dans la lipidation de la protéine ATG8.
La lipidation des protéines de la famille ATG8 se produit probablement aux extrémités fortement incurvées du phagophore.
Cette localisation explique :
- leur fonction dans l'allongement de la membrane,
- la facilitation de leur interaction avec les récepteurs liés au cargos lors de la biogenèse de l'autophagosome de novo pendant l'autophagosome sélectif.
Fermeture du phagophore
Biologie cellulaire et moléculaireConstituants de la celluleSystème endomembranaireRéticulum endoplasmiqueAppareil de GolgiEndosomesLysosomesPeroxysomesAutophagieMacroautophagieMicroautophagieAutophagie chaperonnes (CMA)Autophagie non canoniqueProtéines ATGSystème UPS (Ubiquitine-Protéasome)Transport membranaireTrafic vésiculaireFusion/fission membranaireEndocytoseVoie sécrétoireMoteurs moléculairesVoies de signalisation