


Autophagie
Macroautophagie
Formation de l'autophagosome
Initiation du phagophore
Translocation et activation du complexe ULK/Atg1
- Vue d'ensemble du système endomembranaire
- Autophagie
- Système UPS (Système UPS (Ubiquitine-protéasome))
- Fusion et fission membranaire
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

Le complexe ULK/Atg1 doit subir une translocation et une activation au niveau des sites de nucléation du phagophore pour déclencher la macroautophagie.
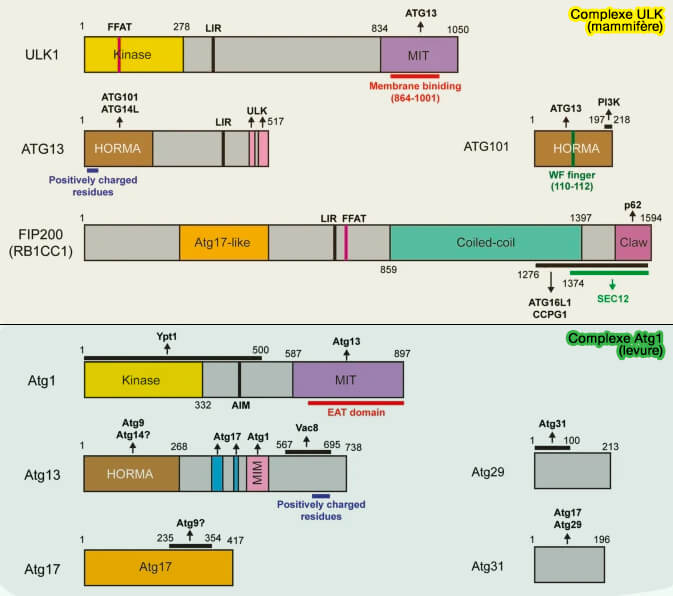
(Figure : vetopsy.fr d'après Nishimura et Tooze)
Translocation du complexe ULK/Atg1
On ne comprend pas bien ce qui provoque la translocation du complexe ULK en sites de nucléation de phagophore lors de dénutrition ( attache membranaire d'ULK).
attache membranaire d'ULK).
Rôles d'ATG9
1. Dans des conditions riches en nutriments, l'ATG9A est principalement située dans la région périnucléaire :
-
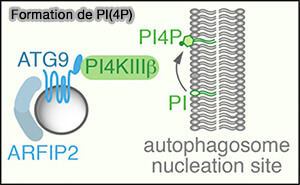
Formation de PI(4)P
(Figure : vetopsy.fr d'après Judith et coll) - mais aussi, moins fortement, au niveau des endosomes précoces et de recyclage (Atg9A trafficking through the recycling endosomes 2016).
2. Pendant la dénutrition, l'ATG9A périnucléaire diminue concomitamment avec une augmentation d'une population vésiculaire de 30–60 nm riche en arfaptines et en PI4-kinase, PI4KIIIβ, kinase qui produit du PI(4)P, au niveau des omégasomes (ATG9A shapes the forming autophagosome through Arfaptin 2 and phosphatidylinositol 4-kinase IIIβ 2019).

ATG9 et ses rôles sont étudiés dans un chapitre spécial.
3. Le trafic d'ATG9A dépend du complexe ULK, bien qu'une analyse plus approfondie soit nécessaire pour confirmer si ATG9A interagit directement avec le complexe ULK (Regulation of mATG9 trafficking by Src- and ULK1-mediated phosphorylation in basal and starvation-induced autophagy 2016).
Chez la levure, l'extrémité N-terminale d'Atg9 se lie également au domaine N-terminal HORMA d'Atg13, ce qui facilite le recrutement des vésicules Atg9 au PAS (Atg13 HORMA domain recruits Atg9 vesicles during autophagosome formation 2015).
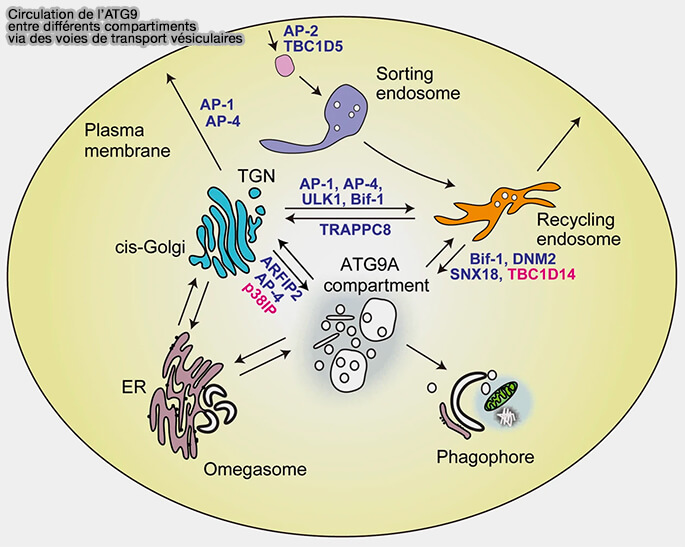
(Figure : vetopsy.fr d'après Nishimura et Tooze)
Rôles de COPII
L'étape d'initiation a également été spatialement liée aux vésicules COPII, qui servent de transporteurs réticulum endoplasmique (RE) ➞ appareil de Golgi.
1. Dans les cellules de mammifères, SEC12, l'activateur de l'assemblage COPII, est associé à l'extrémité C-terminale de FIP200.
2. Toutefois, cette interaction est principalement nécessaire dans le remodelage des sites de sortie du RE (ERES), régions du RE spécialisées dans la formation de vésicules COPII (Remodeling of ER‐exit sites initiates a membrane supply pathway for autophagosome biogenesis 2017).
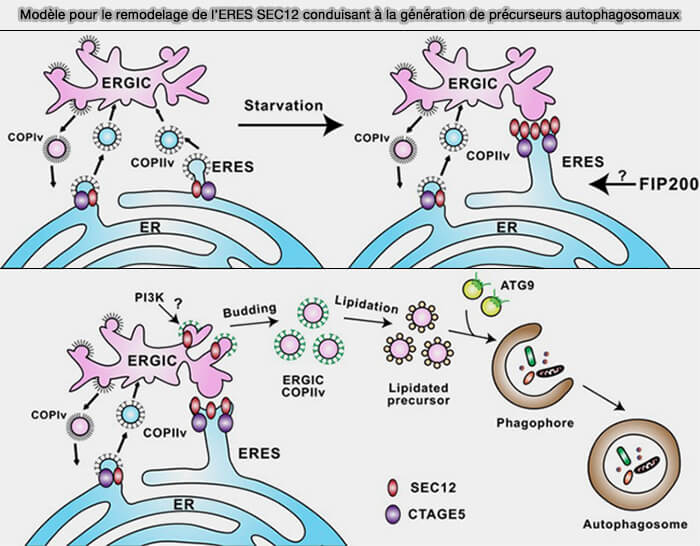
(Figure : vetopsy.fr d'après Ge et coll)
Rôles des VAPA/B du RE
ULK1 et FIP200 interagissent avec les protéines intégrales du RE, les protéines VAP (VAPA et VAPB), qui peuvent attacher le RE à d'autres membranes cellulaires au niveau des sites de contact membranaire (The ER Contact Proteins VAPA/B Interact with Multiple Autophagy Proteins to Modulate Autophagosome Biogenesis 2018).
- Les protéines VAP se lient directement aux protéines à motif FFAT (deux phénylalanines FF in Acidic Tract).
- ULK1 et FIP200 ont des motifs fonctionnels FFAT, suggérant que l'association membranaire de l'ULK1 et du FIP200 est régulée par les protéines VAP.
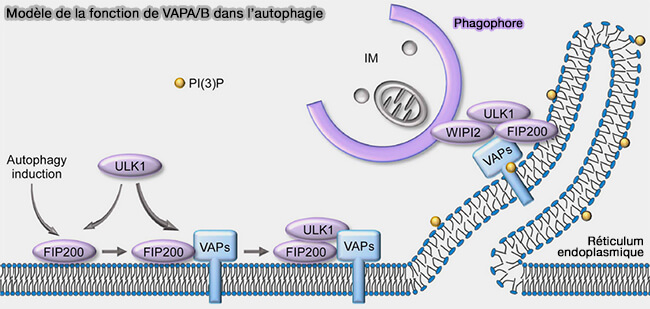
(Figure : vetopsy.fr d'après Zhao et coll)
Ainsi, Ypt1/Rab1, SEC12 et les VAP faciliteraient l'initiation de l'autophagie par le recrutement des composants du complexe ULK.

L'inhibition de ces interactions provoque un déficit partiel de l'autophagie.
Rôles des sous-domaines PIS
Le complexe ULK est localisé en sous-domaines environnementaux enrichis en phosphatidylinositol synthase (PIS).
Le RE est le site de la plupart des synthèses lipidiques dans la cellule, et plusieurs études ont suggéré que la biogenèse de l'autophagosome peut être localisée dans des régions du RE où les enzymes de synthèse lipidique sont concentrées.
1. PIS1 colocalise avec les marqueurs d'autophagie précoces clés, y compris ULK1 et FIP200 (Autophagosome formation is initiated at phosphatidylinositol synthase‐enriched ER subdomains 2017).
2. PIS1 colocalise plus largement avec la protéine VMP1 (vacuole membrane protein 1), protéine des sites de contact (MCS), trouvée à la fois dans les autophagosomes et les endosomes, et colocalisée avec l'enzyme choline/éthanolamine phosphotransférase CEPT1 (Vacuole membrane protein 1 marks endoplasmic reticulum subdomains enriched in phospholipid synthesizing enzymes and is required for phosphoinositide distribution 2018).
- VMP1 interagit également avec la protéine transmembranaire TMEM41B localisée au RE, qui joue un rôle dans la mobilisation des lipides (CRISPR screening using an expanded toolkit of autophagy reporters identifies TMEM41B as a novel autophagy factor 2019).
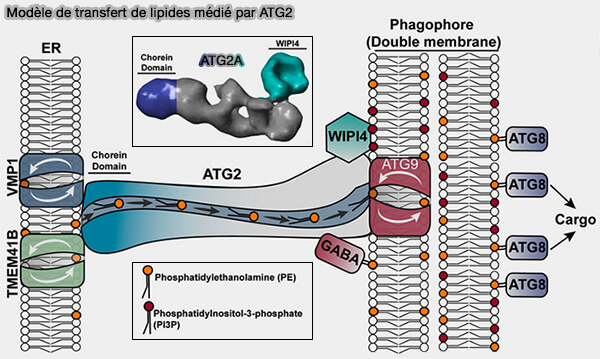
(Figure : vetopsy.fr d'après MacEwan et coll)
- Les KO de TMEM41B, VMP1 et ATG2 bloquent la biogenèse de l'autophagosome à un stade précoce cohérent avec un échec de l'expansion ( extrusion directe des lipides lors de l'élongation du phagophore).
Activation du complexe ULK
La phosphorylation de ULK est la clé de sa régulation (Activation of autophagy depends on Atg1/Ulk1-mediated phosphorylation and inhibition of the Hsp90 chaperone machinery 2023).

Vous pouvez lire : Structural Basis of Autophagy Regulatory Proteins (2019), qui développe les structures et la régulation de mTORC et d'AMP dans la macroautophagie.
(Figure : vetopsy.fr d'après Melia et coll)
1. La phosphorylation d'ULK peut s'effectuer (Role of AMPK-mTOR-Ulk1/2 in the Regulation of Autophagy: Cross Talk, Shortcuts, and Feedbacks 2012) :
- par AMPK (AMP-activated protein kinase), senseur du niveau énergétique (The lysosome: A crucial hub for AMPK and mTORC1 signalling 2017),
- par autophosphorylation, en particulier de sa boucle d'activation loop, i.e. Thr226 dans Atg1 (Autophosphorylation Within the Atg1 Activation Loop Is Required for Both Kinase Activity and the Induction of Autophagy in Saccharomyces cerevisiae 2010).
-
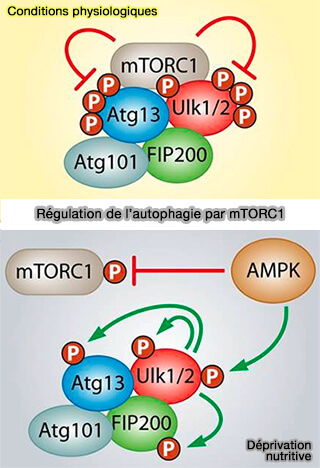
Régulation de l'autophagie par mTORC1
(Figure : vetopsy.fr d'après Alers et coll)
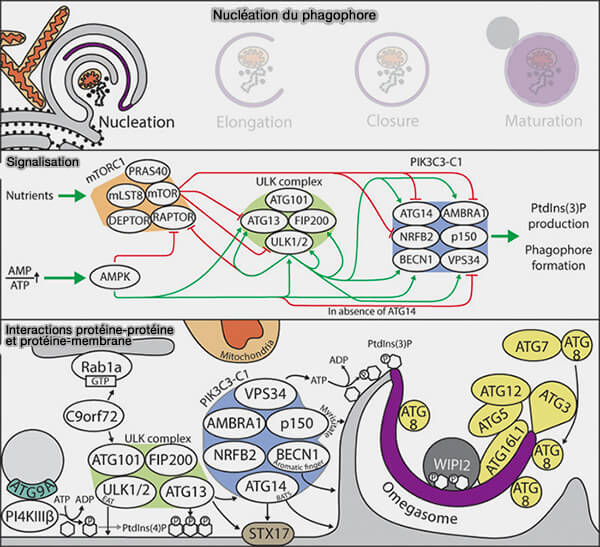
a. ULK1/2 joue alors un rôle prépondérant dans la phosphorylation d’ATG13 et FIP200, entraînant leur activation, ainsi que celle de nombreux substrats comme Atg2 et Atg9 ( tableau).
AMPK et ULK1 phosphorylent Raptor de mTORC1 pour l'inhiber.
Remarque : les phosphorylations sont résumées dans un tableau.
b. Ces phosphorylations conduisent au recrutement de PIK3C3-CI et de la production de PI(3)P, ce qui entraîne la formation d'omégasomes PI(3)P-positifs qui fonctionnent comme plates-formes d'élongation par le phagophore.
Remarque : En plus de cette phosphorégulation positive de l'activité ULK1, l'ubiquitination de LYS-63 de l'ULK1 par l'E3-ligase TRAF6 liée à AMBRA1 favorise la stabilisation et la fonction des complexes ULK1.
c. Une étude récente a montré que Hsp90 se lie et forme un complexe avec Atg1/Ulk1, qui supprime son activité kinase (Activation of autophagy depends on Atg1/Ulk1-mediated phosphorylation and inhibition of the Hsp90 chaperone machinery 2023).
- L’activation d’Atg1/Ulk1 provoque la phosphorylation de Hsp90, inhibant son activité de l’ATPase et modifiant la dynamique du chaperon.
- Ces événements conduisent à la dissociation du complexe Hsp90/Atg1/Ulk1 et à l’activation de Atg1/Ulk1, essentielle à l’initiation de l’autophagie.
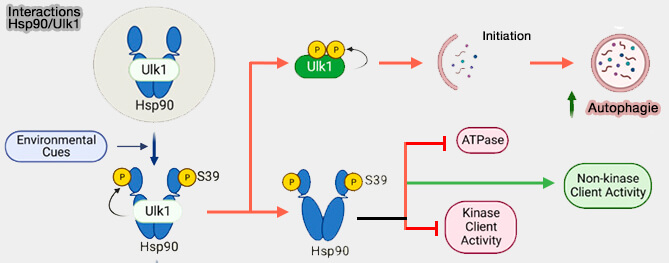
(Figure : vetopsy.fr d'après Backe et coll)
Remarque : Hsp90 est aussi un cochaperon de Hsp70 dans l'autophagie médiée par des protéines chaperonnes (CMA).
2. En revanche, dans des conditions nutritives physiologiques, mTOR (mammalian target of rapamycin), véritable senseur des variations en nutriments au sein de la cellule, vient inhiber l’activité du complexe ULK.
- mTOR phosphoryle ULK1/2 et ATG13 par le biais du complexe mTORC1 ou mTOR complexe 1 (Regulation of the mTOR complex 1 pathway by nutrients, growth factors, and stress 2010).
- Dès lors, la dissociation de mTORC1 du complexe d’initiation, notamment suite à une déprivation nutritive, entraîne son activation (mTOR inhibits autophagy by controlling ULK1 ubiquitylation, self-association and function through AMBRA1 and TRAF6 2013).
Nucléation du phagophore
Biologie cellulaire et moléculaireConstituants de la celluleSystème endomembranaireRéticulum endoplasmiqueAppareil de GolgiEndosomesLysosomesPeroxysomesAutophagieMacroautophagieMicroautophagieAutophagie chaperonnes (CMA)Autophagie non canoniqueProtéines ATGSystème UPS (Ubiquitine-Protéasome)Transport membranaireTrafic vésiculaireFusion/fission membranaireEndocytoseVoie sécrétoireMoteurs moléculairesVoies de signalisation