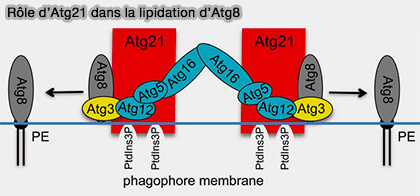
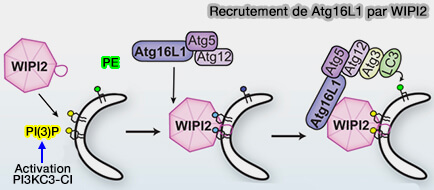
Les protéines Atg18/WIPI appartiennent à une grande famille de protéines appelées PROPPIN (β-PROPellers that bind PIs), que l'on trouve chez la plupart des eucaryotes.
Vue d'ensemble des protéines PROPPIN
1. La famille des protéines PROPPIN sont une famille de protéines d'autophagie conservée dans l'ensemble des eucaryotes et qui se lient aux phosphoinositides, d'où leur nom.
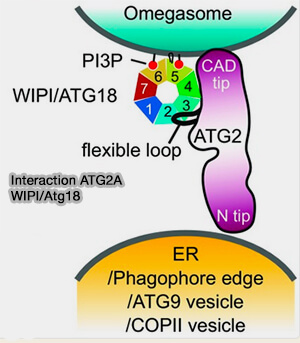
Interaction ATG2A WIPI/Atg18
(Figure : vetopsy.fr
d'après Chowdhury et coll)
Malgré les similitudes entre Atg18 et les WIPI,on ne sait pas encore si les protéines WIPI régulent les niveaux de PI(3,5)P2 dans les cellules de mammifères.
WIPI-2 n'a pas été testé jusqu'à présent en raison de son instabilité.
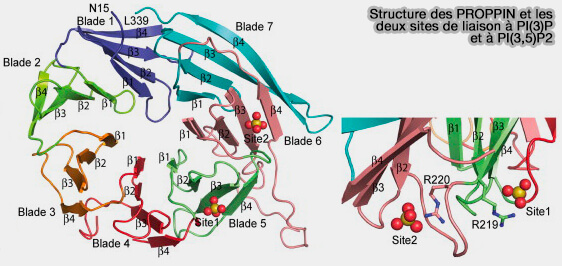
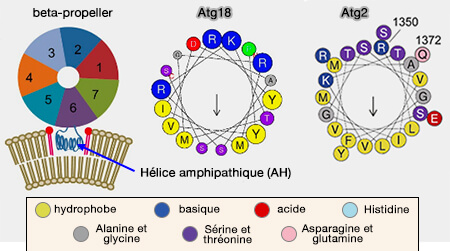
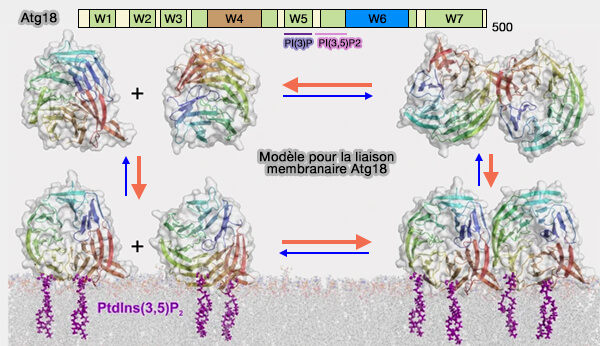
L'association membranaire directe des protéines Atg18/WIPI est contrôlée par l'hélice et les deux sites de liaison.
2. Le processus de dimérisation des WIPI est encore obscur.
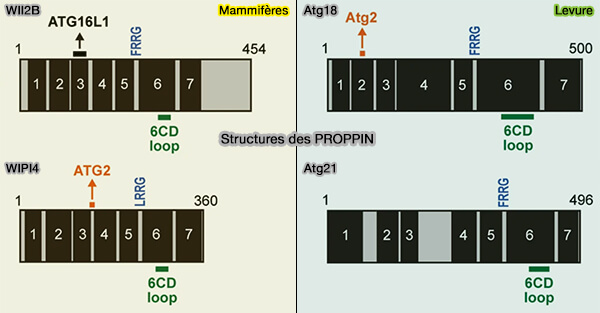
3. Comme les sites de liaison avec l'Atg2/ATG2 par WIPI4 et de liaison Atg16L1 par WIPI2 sont situés à la surface opposée du motif FRRG, cette architecture permet aux Atg18/WIPI d'interagir simultanément avec les protéines ATG en aval.
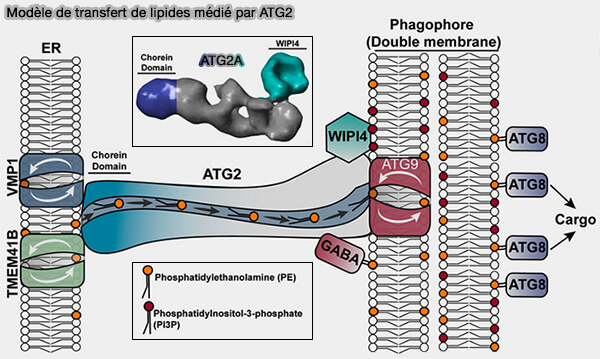
Modèle de transfert de lipides médié par ATG2
(Figure : vetopsy.fr d'après MacEwan et coll)
Remarque : ATG18 peut avoir un rôle distinct dans la fission de la membrane vacuolaire ( PROPPIN et fission lysosomale).
Rôles des WIPPI
Chez la levure
1. Atg18 est essentiel pour l'autophagie, mais aussi pour d'autres fonctions.
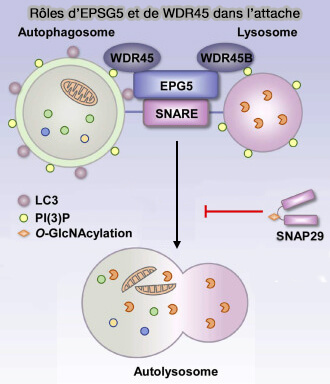
Rôles d’EPSG5 et de WDR45 dans l’attache
(Figure : vetopsy.fr d'après Ji et coll)
a. Atg18 forme un complexe avec Atg2, et la formation de ce complexe est nécessaire pour le recrutement des deux protéines aux phagophores ( liaisons membranaires).
b. Atg18 intervient dans d'autres régulations comme celle :
Contrairement aux PROPPIN de levure, la fonction de chaque protéine WIPI et les relations fonctionnelles entre WIPI1-4 n'ont pas été complètement élucidées.
Si le rôle de WIPI2 est bien établi, les rôles de WIPI1, WIPI3 et WIPI4 dans l'autophagie restent mal compris, et il n'est toujours pas clair si les protéines WIPI contribuent au recrutement d'ATG2A/B dans les cellules de mammifères.
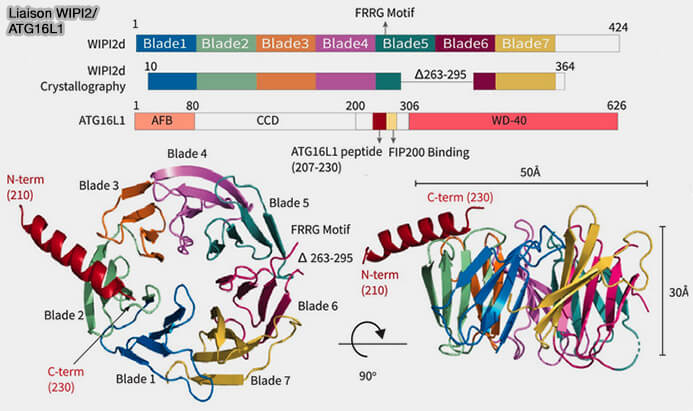
La région d’interaction WIPI2 (W2IR) d'ATG16L1 comprenant les résidus 207 à 230 adopte une conformation hélicoïdale alpha et se lie dans un sillon électropositif et hydrophobe entre les pales 2 et 3 de l’hélice WIPI2β.
La mutation des résidus à l’interface réduit ou bloque le recrutement d’Atg12/ATG5/ATG16L1 et la conjugaison de la protéine ATG8 LC3B aux membranes synthétiques. Les mutants d’interface montrent une diminution de l’autophagie induite par la famine.
Ces résidus ne sont pas conservés dans ATG16L2 qui n'a aucun rôle dans l'autophagie.
4. Bien que WIPI3, WIPI4 et ATG2A/B soient exprimés de manière ubiquitaire, la délétion simultanée de WIPI3 et de WIPI4 provoque un défaut d'autophagie observé uniquement dans les cellules neuronales cultivées et les cerveaux de souris, mais pas dans les cellules non neuronales cultivées.
L'interaction entre ATG2A/B et WIPI4 (et probablement WIPI3) ne serait pas indispensable pour le flux autophagique.
Par contre, ATG2A/B seraient recrutées dans des phagophores par interaction entre les protéines de la famille d'ATG8 telles que GABARAP, GABARAPL1 et LC3A.
Les mutations dans le WDR45, qui codent pour WIPI4, provoquent la neurodegeneration with brain iron accumulation 5, qui se caractérise généralement par des symptômes neurodéveloppementaux statiques au stade précoce de la maladie et de la neurodégénérescence à un stade avancé de la maladie,.
Les mutations dans WDR45B, qui codent WIPI3, provoquent également un trouble neurodéveloppemental complexe appelé syndrome d'El-Hattab-Alkuray.
Les mutations de WIPI2 provoquent un trouble neurodéveloppemental accompagné d'anomalies osseuses et cardiaques (Oocyte/zygote/embryo maturation arrest 6).






 PROPPIN et fission lysosomale).
PROPPIN et fission lysosomale).