Les LRO (Lysosome-Related Organelles ou organites liés aux lysosomes) comprennent un groupe diversifié d'organites sub-cellulaires spécifiques à un type de cellule et liées à la membrane qui dérivent au moins en partie du système endo-lysosomal, mais qui ont un contenu, des morphologies et des fonctions uniques pour soutenir des rôles physiologiques dans des cellules spécifiques.
2. Les LRO englobent un large éventail de structures de morphologies et de fonctions distinctes qui sont spécifiques de la physiologie de leur cellule hôte, i.e. leur contenu est trié au cours de la naissance des LRO (Lysosome-related organelles: Unusual compartments become mainstream 2014).
3. De plus, la plupart (sinon la totalité) des LRO sont des organites sécrétoires régulés, et la sécrétion au bon moment est essentielle pour leur fonction cellulaire.
Par exemple, le contenu :
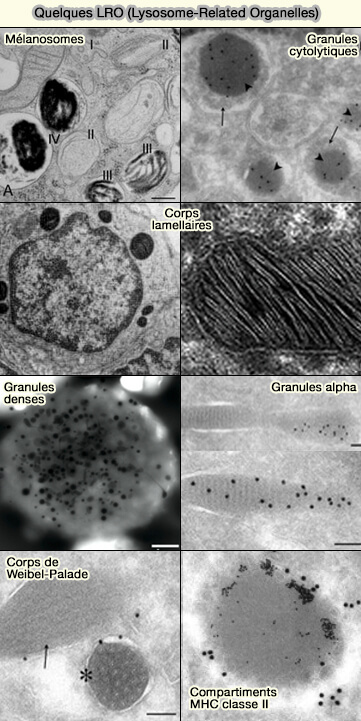
des granules cytolytiques ne doit être dirigé vers une cellule cible qu'après la reconnaissance immunitaire,
des granules alpha et des granules denses ne doit être libéré que lors de l'activation des plaquettes sur les sites de lésions vasculaires.
Une régulation appropriée de la biogenèse, le maintien/la protection contre la dégradation et la sécrétion de LRO sont nécessaires pour une fonction physiologique normale.
Les différents LRO
Liste des LRO
La liste des LRO est visible dans un chapitre spécial.
Petit aparté sur les mélanosomes
Pourquoi ce petit aparté sur les mélanosomes ?
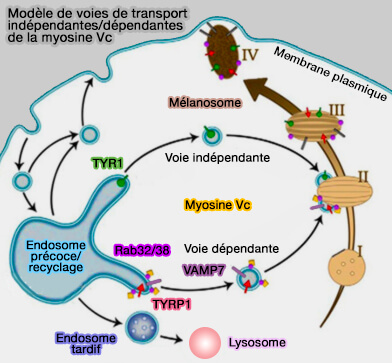
1. Les mélanosomes, contenus dans les mélanocytes, sont les LRO les plus étudiés, i.e. de nombreux travaux sont basés sur les molécules intervenant dans leur trafic, comme :
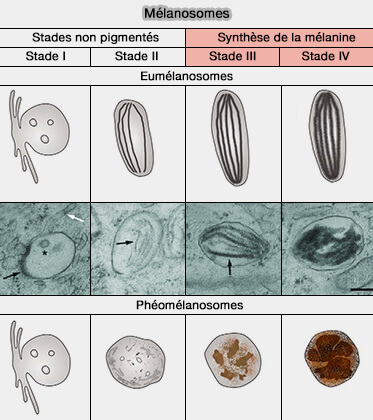
b. Les mélanosomes de stade II contiennent des fibrilles complètement formées qui font adopter au mélanosome une forme ellipsoïdale et servent également d'échafaudage pour la pigmentation de la mélanine.
c. Les mélanosomes de stade III et IV voient la synthèse de la mélanine qui apparaît dense aux électrons et qui se dépose progressivement sur les fibrilles jusqu'à ce que la structure interne du mélanosome soit complètement obscurcie à la fin du stade IV.
Contrairement aux eumélanosomes, les phéomélanosomes sont toujours sphériques au cours du développement et ne contiennent que de la matière protéique amorphe et de la mélanine granulaire, mais jamais de filaments protéiques clairs ni de matrice.
Ils fusionnent par la suite avec des membranes endosomales portant CD63 et la P-sélectine.
b. Par contre, les granules denses ont longtemps été considérés comme des granules sécrétoires classiques, mais présentent certaines caractéristiques des LRO.
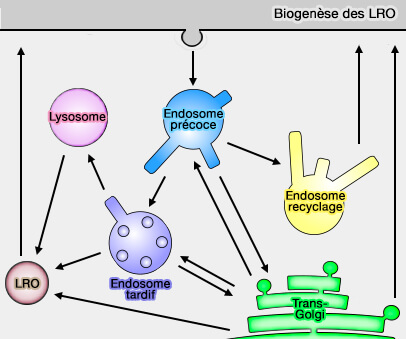
2. Dans d'autres types de cellules qui utilisent des LRO, tels que les mélanocytes et les cellules endothéliales, des lysosomes conventionnels existent également, et la biogenèse des LRO diverge de la voie utilisée par les lysosomes conventionnels.
Les protéines destinées à la sécrétion sont ciblées vers les LRO pour la voie de sécrétion spécialisée.
Biogenèse et trafic des LRO
1. La biogenèse et le trafic des LRO nécessitent de nombreuses molécules et complexes, comme, de manière non exhaustive :
Biogenèse des LRO
(Figure : vetopsy.fr)
l'adaptine AP-3 et quelquefois AP-1,