de tube, de pont et de tunnelmultimériques ou monomériques, ce qui a l'énorme avantage de déplacer le lipide dit par glissement (sliding en anglais) sans ou avec un minimum de mouvements des protéines, i.e. ce qui permet une augmentation substantielle de la vitesse du transfert.
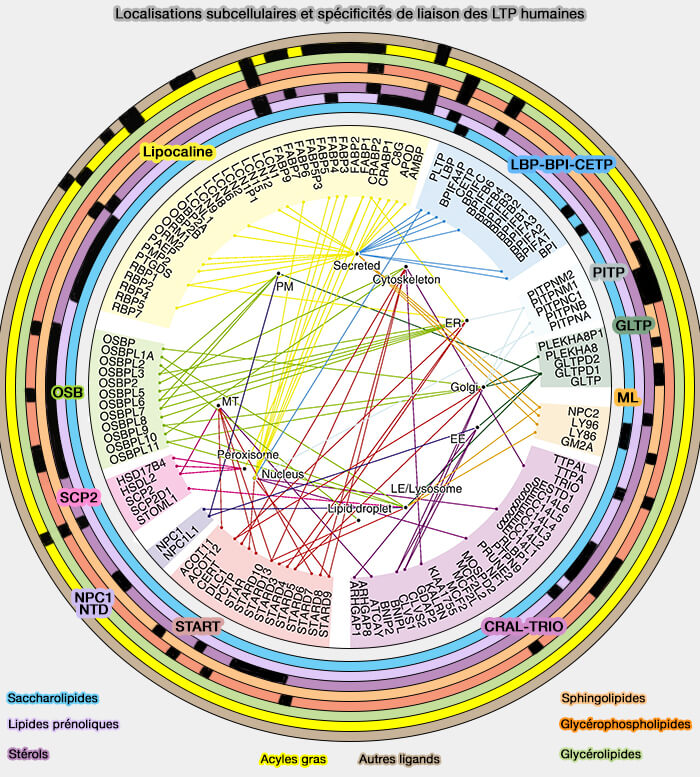
Localisations subcellulaires et spécificités de liaison des LTP humaines
(Figure : vetopsy.fr d'après Chiapparino et coll)
Vue d'ensemble des transferts lipidiques par pont ou tunnel
1. Les LTP en forme de ponts ou tunnels protéiques comme Vps13 ou Atg2 sont des protéines beaucoup plus grandes 1 500 à 3 000 résidus, par rapport aux LTP tubulaires (TULIP).
Remarque : cette classification peut être modifiée selon les auteurs, i.e. les TULIP ou les LTP par exemple, peuvent être considérés comme des ponts entre les membranes.
2. Alors que les navettes en forme de boîte ont un point d'accès unique à leurs cavités de liaison, les LTP en forme de pont s'ouvrent de chaque côté de la membrane.
Ce long pont hydrophobe permet aux queues lipidiques hydrophobes de coulisser au travers tandis que la protéine reste stationnaire.
LTP bactériennes
Domaine MCE de MlaD et héxamérisation en disque
(Figure : vetopsy.fr d'après Ekiertet coll)
1. Les domaines MCE dans ces complexes se multimérisent.
Tout d'abord, ils forment un disque avec un pore central bordé de résidus hydrophobes par héxamérisation.
Ensuite, les disques s'empilent pour étendre les pores dans un tube hydrophobe.
2. Les protéines MlaD, YebT et PqiB sont des protéines MCE présentes dans l'espace intermembranaire des bactéries.
MlaD est formé par un seul disque qui accepte les lipides transporté par Mlac, une LTP soluble.
PqiB possède 3 disques avec une superhélice en six faisceaux qui forme un pore hydrophobe central.
YetT contient 7 disques empilés qui forme le canal hydrophobe.
Modèle pour le transport des phospholipides médié par MCE à travers le périplasme chez E. coli
(Figure : vetopsy.fr d'après Ekiertet coll)
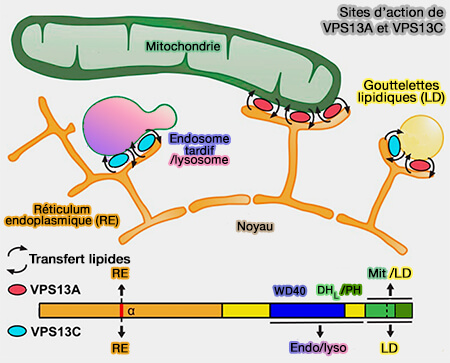
Protéines à domaine choréine : VPS13 et ATG2
Certains processus cellulaires tels que la biogenèse mitochondriale et l’autophagie peuvent nécessiter la production de grandes quantités de membranes biologiques à des échelles physiques relativement grandes et sur des délais courts
1. Des protéines telles que VPS13 ou ATG2 peuvent relier deux membranes par un long pont protéique en forme de tige avec une rainure hydrophobe à travers laquelle les lipides peuvent glisser alors que la protéine reste stationnaire.
Ce mécanisme est parfaitement adapté au transport en vrac des lipides bicouches pour favoriser la croissance membranaire.
2. Ces protéines appartiennent à la famille des protéines à domaine choréine, appelé ainsi car le VPS13A humain est aussi appelé choréine.
L'Atg2 et son rôle dans l'autophagie est étudiée dans un chapitre spécifique.
Structures de Vps13 et Atg2
(Figure : vetopsy.fr d'après plusieurs sources)
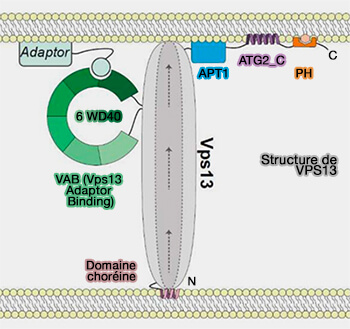
VPS13
Les protéines de la famille VPS13 sont de grosses molécules (300–500 kDa).
Elles sont allongées (200 Å ou plus), i.e. suffisamment pour s'étendre entre les organites au niveau des sites de contact membranaires (MCS).
Elles présentent une rainure hydrophobe sur leur longueur pouvant héberger les glycérolipides qui peuvent ainsi coulisser au travers grâce à leurs queues lipidiques hydrophobes.
Comme la largeur de la rainure est prévue pour accueillir un seul lipide à son point le plus étroit, la structure prédit un transfert unidirectionnel de lipides d'une membrane à une autre.
Domaines de ciblage de VPS13
En aval du domaine de transfert lipidique se trouvent d'autres domaines qui permettent de cibler la localisation et la fonction cellulaire de la protéine.
VAB et adaptateurs
L'association membranaire et la localisation cellulaire sont déterminées par une combinaison de :
motifs structurels présents dans chaque protéine,
d'interactions avec des protéines adaptatrices spécifiques à la membrane se liant aux extrémités C-terminale et N-terminale de la protéine.
Structure générale de VPS13
(Figure : vetopsy.fr d'après Dziurdzik et coll)
2. Toutefois, les 4 protéines VPS13 humaines ont divergé pour se localiser dans différents sous-ensembles de MCS et elles doivent utiliser des déterminants de localisation différents.
Régulation des domaines APT1
(Figure : vetopsy.fr d'après s Kolokowski et coll)
Hélice AH de ATG2A
(Figure : vetopsy.fr d'après Tamura et coll)
2. Le domaine ATG2_C, d'environ 70 à 100 résidus, correspondant à un court segment de la région C-terminale de Atg2, est constitué de deux ou trois hélices dont l'une est amphipathique (AH), ce qui qui permet son insertion dans les membranes, la détection des régions de courbure membranaire et même la déformation membranaire ( hélices amphipatiques).












 VPS13D dans les MCS)
VPS13D dans les MCS)