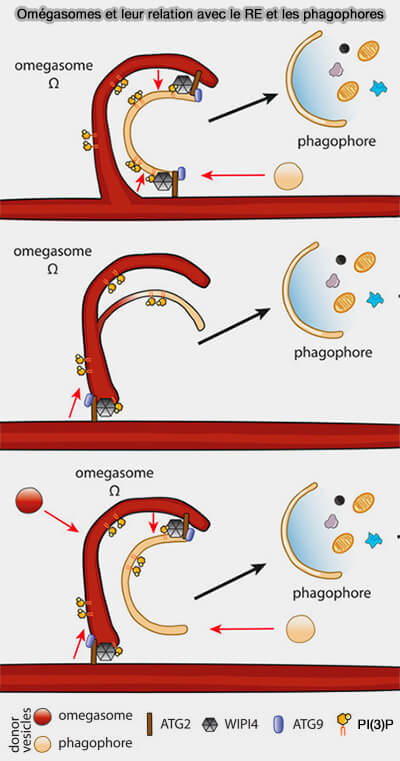
L’initiation du phagophore a lieu au niveau des omégasomes, sous-domaines du réticulum endoplasmique riches en PI(3)P, et le complexe ULK/Atg1 est l'un des premiers facteurs à cibler ces structures membranaires.
La biogenèse des autophagosomes implique plusieurs processus :
1. L’origine des membranes vésiculaires est encore en cours d'études.
a. Chez la levure, cet évènement se déroule au niveau d’un site particulier appelé structure pré-autophagosomale ou site d’assemblage du phagophore (PAS).
2. La formation des omégasomes se produit en 3 minutes et celle des autophagosomes en 10 minutes.
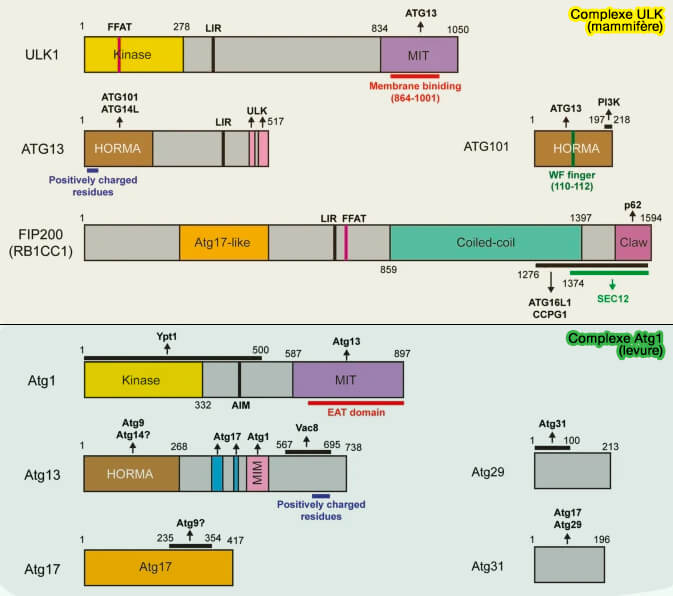
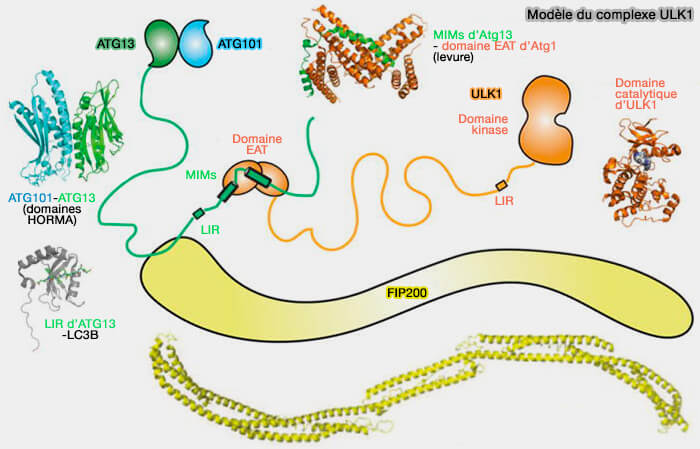
Composition du complexe ULK/Atg1
Le complexe ULK/Atg1 est l'un des premiers facteurs à cibler les structures membranaires au site de formation du phagophore, et son recrutement est indépendant :
Ces interactions ont été proposées pour être impliquées dans la maturation des autophagosomes et/ou une rétroaction négative en dégradant le complexe ULK/Atg1 via l'autophagie.
Complexe Atg1 de la levure
Sructure des composants du complexe ULK1/Atg1
(Figure : vetopsy.fr d'après Nishimura et Tooze)
1. Le complexe Atg1 de la levure est le pendant du complexe ULK
Atg17 intervient dans l'autophagie en vrac, i.e. bulk macroautophagy, tandis qu'Atg11, qui a un rôle similaire et qui peut être considérée comme une autre sous-unité du complexe, est impliquée dans l'autophagie sélective ( autophagie sélective chez la levure).
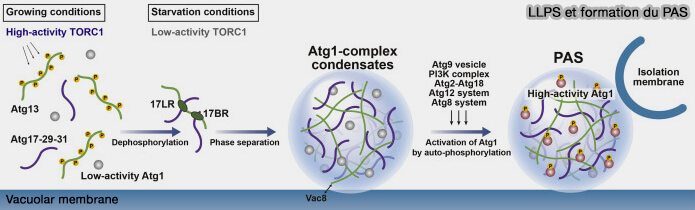
a. Atg13 est déphosphorylée par les phosphatases, ce qui est suffisant pour lier Atg17 et provoque la séparation de phase du complexe Atg1.
b. L’équilibre entre la phosphorylation d’Atg13 par Atg1 et la déphosphorylation par les phosphatases peut être important pour le maintien du PAS de type liquide, qui recrute dynamiquement les protéines ATG au fur et à mesure que l’autophagie progresse.
LLPS et formation du PAS
(Figure : vetopsy.fr d'après Fujioka et Noda)
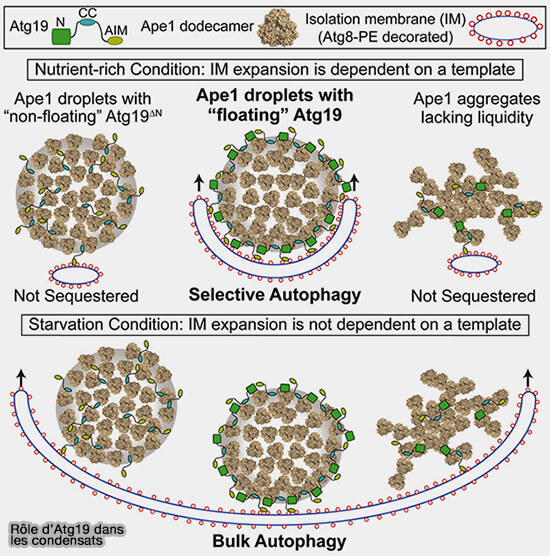
3. Dans des conditions riches en nutriments, Ape1, l’aminopeptidase vacuolaire, est délivrée à la vacuole par autophagie sélective, dans un processus connu sous le nom de Cvt (Cytoplasm-to-vacuole targeting).
La voie de ciblage cytoplasmique vers la vacuole (Cvt) est un type d'autophagie sélective chez Saccharomyces cerevisiae, qui transporte de manière constitutive et sélective les hydrolases vacuolaires vers les vacuoles par autophagie (Structural Biology of the Cvt Pathway 2017).
b. La " flottabilité " d'Atg19, protéine réceptrice spécifique d’Ape1, empêche sa pénétration dans les gouttelettes et se localise à la surface des gouttelettes d’Ape1.
Atg19 régule la taille du complexe Ape1 et cible plusieurs cargos.
Atg19, Atg11 et Atg8-PE lipidé sont nécessaires et suffisants pour la séquestration sélective des gouttelettes d’Ape1 par les membranes et pour former la vésicule Cvt.
c. Cette séquestration est altérée par la solidification mutationnelle des gouttelettes d’Ape1 ou la diminution de la capacité de flottaison d’Atg19
LLPS et formation du PAS
(Figure : vetopsy.fr d'après Yamasaki et coll)
Les condensats, i.e. le PAS pendant l’autophagie induite par la famine et les condensats Ape1 dans la voie Cvt, jouent un rôle essentiel dans l’activation d’Atg1.





 autophagie sélective chez la levure).
autophagie sélective chez la levure).