


Système endo-lysosomal
Trafic des endosomes/lysosomes
Trafic rétrograde (centripète) lié aux microtubules
- Vue d'ensemble du système endomembranaire
- Endosomes
- Lysosomes
- Trafic des endosomes/lysosomes
- Fusion et fission membranaire
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

La distribution spatio-temporelle des organites endo-lysosomaux dépend du transport entraîné par des moteurs de microtubules tels que les kinésines et la dynéine, et par des moteurs de myosine à base d'actine.
Les organites se déplacent dans tout le cytoplasme le long du cytosquelette, i.e. microtubules et filaments d'actine, et ces mouvements sont finement régulés ( vue d'ensemble du trafic des endosomes).
vue d'ensemble du trafic des endosomes).
1. Les protéines motrices de microtubules transportent les organites rapidement sur de grandes distances et de manière bidirectionnelle.
- La plupart des kinésines déplacent les organites de l'extrémité moins vers l'extrémité plus (transport antérograde ou centrifuge).
- La dynéine provoque le déplacement en sens inverse, de l'extrémité plus vers l'extrémité moins (transport rétrograde ou centripète).
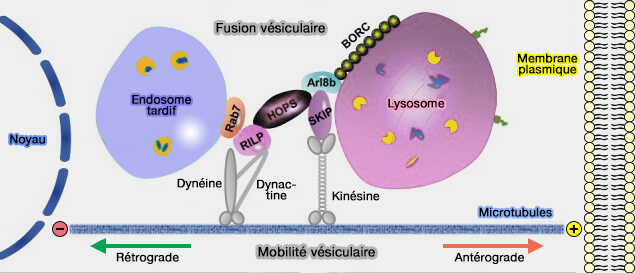
(Figure : vetopsy.fr d'après Khattern et coll)
2. Les moteurs de l'actine, i.e. les myosines, interviennent sur de courtes distances et de manière lente (transport lié au cytosquelette d'actine).
3. Le réticulum endoplasmique (RE), omniprésent dans tout le cytoplasme, joue un rôle majeur dans le contrôle du mouvement et du positionnement global des organites endo-lysosomaux ( trafic des endosomes lié au MCS du RE).
Généralités sur le transport rétrograde (centripète) lié aux microtubules
Le transport rétrograde (centripète) utilise :
- la dynéine comme protéine motrice,
- les sites de contact membranaire (MCS) réticulum endoplasmique/endosomes.
(Figure : vetopsy.fr d'après Lee et coll)
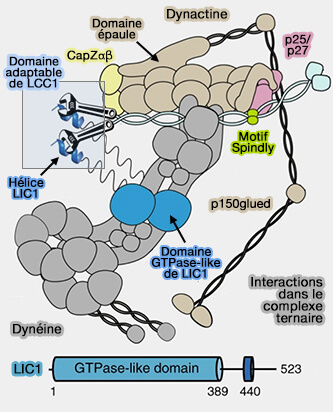
1. La dynéine, contrairement aux kinésines, ne possède qu'une seule chaîne lourde, mais se présente sous forme de dimère.
La diversification structurelle et fonctionnelle de la dynéine est obtenue par l'association de la chaîne lourde avec :
- diverses chaînes intermédiaires (IC), légères (LC) et intermédiaires légères (LIC),
- de nombreux adaptateurs de cargos.
2. Un autre complexe multi-sous-unité nommé dynactine s'associe à la dynéine pour activer son transport vers les extrémités pointées (-) des microtubules.

La dynéine et ses adaptateurs, la dynactine et les mécanismes d'activation sont étudiés en détail dans des chapitres spéciaux avec les moteurs moléculaires.
3. Pratiquement tous les organites endo-lysosomaux peuvent recruter de la dynéine/dynactine selon un modèle similaire à celui des kinésines ( tableau) :
- une petite GTPase Rab se lie à l'organite cible,
- un effecteur de cette Rab interagit directement ou indirectement avec le moteur.
Transport rétrograde (centripète) des endosomes précoces
Le transport centripète des endosomes précoces, par exemple, est médié par Rab5 et son effecteur FHF.
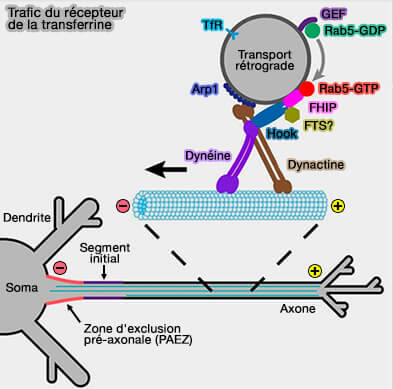
(Figure : vetopsy.fr d'après Guo et coll)
1. FHF (Fused Toes) est composé de sous-unités :
- FHIP,
- Hook - et FTS ? - (Hook is an adapter that coordinates kinesin-3 and dynein cargo attachment on early endosomes 2014) .
2. FHIP est la sous-unité qui interagit directement avec Rab5 (Rab5 and its effector FHF contribute to neuronal polarity through dynein-dependent retrieval of somatodendritic proteins from the axon 2016).
3. Hook se lie simultanément à la dynéine et à la dynactine, i.e. c'est un adaptateur, stabilisant ce complexe et améliorant la processivité du transport (Assembly and activation of dynein–dynactin by the cargo adaptor protein Hook3 2016 et Hook Adaptors Induce Unidirectional Processive Motility by Enhancing the Dynein-Dynactin Interaction 2016).
Transport rétrograde (centripète)
des endosomes tardifs/lysosomes
La dynéine peut être recrutée via :
- Rab7 et RILP, en présence d'un taux de cholestérol élevé,
- ALG2, le capteur de Ca++ cytosolique, et canal calcique lysosomal TRPML1 régulées par PI(3,5)P2 et le calcium,
-
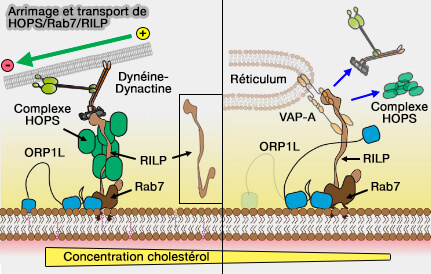
Attache et transport de HOPS/Rab7/RILP par la dynéine
(Figure : vetopsy.fr d'après van der Kant et coll) positionnement des lysosomes régulé par les nutriments).
Rab7/RILP
Vue d'ensemble
Le transport centripète des endosomes tardifs/lysosomes dépend de Rab7 et de son effecteur RILP (The Rab7 effector protein RILP controls lysosomal transport by inducing the recruitment of dynein-dynactin motors 2001).
Rab7/RILP interagit avec :
- la sous-unité de dynactine p150Glued (Activation of endosomal dynein motors by stepwise assembly of Rab7–RILP–p150Glued , ORP1L, and the receptor βlll spectrin 2007),
- la chaîne intermédiaire légère de dynéine (A Ras-like domain in the light intermediate chain bridges the dynein motor to a cargo-binding region 2014).
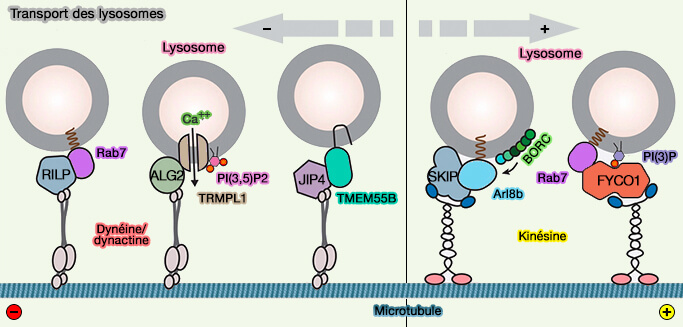
(Figure : vetopsy.fr d'après Cabukusta et Neefjes)
Exemple du cholestérol : ORP1L
Rôle dans le trafic
1. Le cholestérol peut altérer la fluidité membranaire et former des microdomaines lipidiques dans les membranes dans lesquels certaines protéines sont exclues et d'autres sont enrichies (Functional rafts in cell membranes 1997).
-
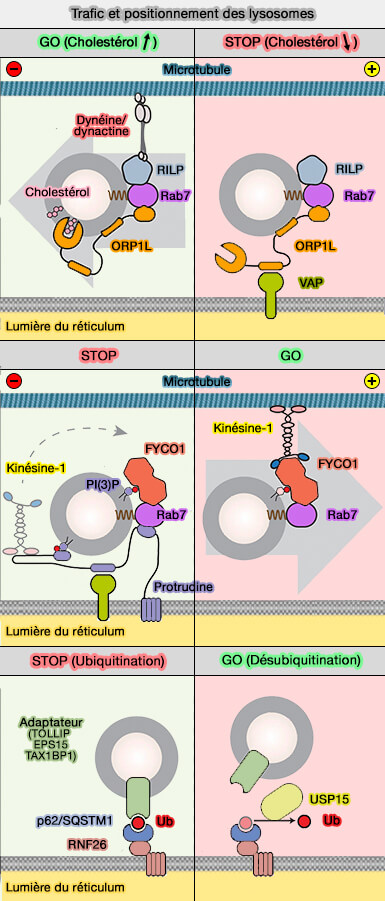
Trafic et positionnement des lysosomes
(Figure : vetopsy.fr d'après Cabukusta et coll) - Le cholestérol peut être impliqué dans la formation de vésicules intraluminales (ILV) par une voie dépendante de ESCRT ou une voie indépendante nécessitant les tétraspanines ou les céramides.
Les sites de contact RE/lysosome sont impliqués dans l'homéostasie du cholestérol (Cholesterol-binding molecules MLN64 and ORP1L mark distinct late endosomes with transporters ABCA3 and NPC1 2013 et Annexin A1 Tethers Membrane Contact Sites that Mediate ER to Endosome Cholesterol Transport 2016 et STARD3 mediates endoplasmic reticulum-to-endosome cholesterol transport at membrane contact sites 2017).
2. Le complexe Rab7-RILP-dynéine-dynactine s'associe à un capteur de cholestérol nommé ORP1L, forme longue de l'ORP1, qui fait partie de la famille des protéines OSBP/ORP/Osh - Oxysterol-binding proteins/OSBP-Related Proteins mammaliennes et OSBP-homolog de la levure - ( échange de lipides dans les MCS RE/endosomes).
a. ORP1L adopte différentes conformations en fonction du taux de cholestérol cellulaire (Cholesterol sensor ORP1L contacts the ER protein VAP to control Rab7–RILP–p150Glued and late endosome positioning 2009 et A Role for Rab7 in the Movement of Secretory Granules in Cytotoxic T Lymphocytes 2011).
b. La maturation correcte des autophagosomes nécessite leur transport par la dynéine vers la région périnucléaire de la cellule ou MTOC (Cholesterol and ORP1L-mediated ER contact sites control autophagosome transport and fusion with the endocytic pathway 2016).
a. À faible taux de cholestérol, un motif FFAT dans ORP1L interagit avec la protéine VAPA (Vamp-Associated Protein) au niveau des sites de contact membranaire (MCS) entre les endosomes tardifs/lysosomes et le réticulum endoplasmique (Activation of endosomal dynein motors by stepwise assembly of Rab7–RILP–p150Glued, ORP1L, and the receptor βlll spectrin 2007 et Late endosomal transport and tethering are coupled processes controlled by RILP and the cholesterol sensor ORP1L 2013).
- L'interaction avec VAPA dissocie la dynéine/dynactine du complexe Rab7-RILP, entraîne une localisation périphérique des endosomes tardifs et empêche le transport des autophagosomes vers le MTOC (4).
- Les sites de contact médiés par ORP1L inhibent également la localisation de PLEKHM1 à Rab7 (2).
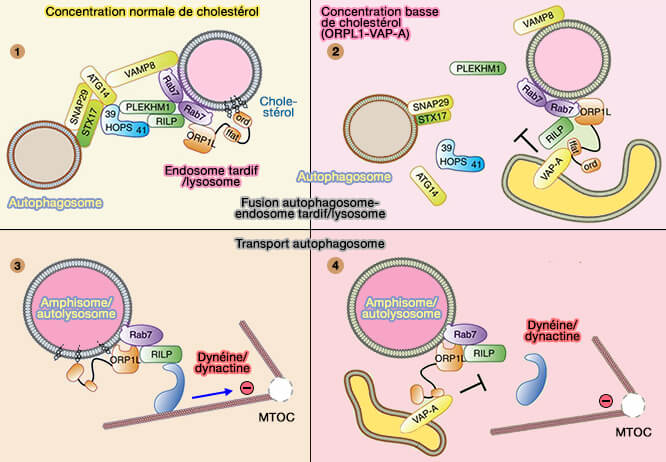
(Figure : vetopsy.fr d'après Wijdeven et coll)
b. Lors de taux de cholestérol adéquat, on assiste à la libération d'ORP1L de VAPA, par l'occlusion du motif FFAT, ce qui permet :
- le recrutement de la dynéine pour le transport des autophagosomes vers le MTOC (3),
- la formation du complexe Rab7-RILP-PLEKHM1 qui recrute le complexe HOPS aux endosomes tardifs/lysosomes afin qu'ils puissent fusionner avec les autophagosomes (1).
Ce mécanisme explique non seulement comment le transport endosome tardif/lysosome est contrôlé par le cholestérol et le réticulum endoplasmique, mais aussi le phénotype de regroupement endosome tardif/lysosome causé par l'accumulation de cholestérol endolysosomal dans les cellules des patients atteints de maladie Niemann-Pick de type C.
Autres rôle de ORP1L
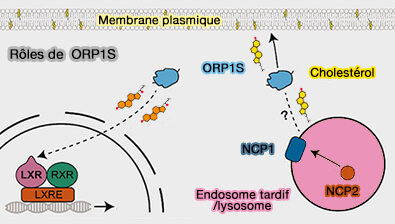
1. ORP1L transporte le cholestérol du réticulum endoplasmique (RE) vers la membrane limitante des endosomes tardifs/lysosomes et dans les vésicules intraluminales, formant les endosomes ou corps multivésiculaires (MVE/MVB).
2. ORP1L transporte le cholestérol des lysosomes, délivré par NPC1 et NPC2, vers le RE ( sortie lysosomale du cholestérol).
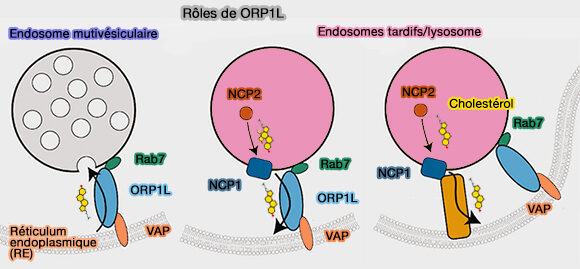
(Figure : vetopsy.fr d'après Aw et coll)
3. ORP1L relie les LEL au RE alors que GRAMD1B transporte le cholestérol LEL vers le RE.
Remarque 1 : la signalisation PI(4)P par ORP1L dans les contacts à trois voies contribuerait à la division mitochondriale (ORP1L mediated PI(4)P signaling at ER-lysosome-mitochondrion three-way contact contributes to mitochondrial division 2021).
(Figure : vetopsy.fr d'après Aw et coll)
Remarque 2 : ORP1S, la forme courte d'ORP1, transporte le cholestérol vers la membrane plasmique et pourrait délivrer des oxystérols aux LXR localisés dans le noyau pour réguler positivement l'expression de l'apolipoprotéine E.
PLEKHM1
PLEKHM1 (Pleckstrin Homology and RUN domain containing M1), de la famille SKIP, se trouve sur les endosomes tardifs, les phagosomes et les autophagosomes (PLEKHM1 Regulates Autophagosome-Lysosome Fusion through HOPS Complex and LC3/GABARAP Proteins 2015 et PLEKHM1 Regulates Salmonella-Containing Vacuole Biogenesis and Infection 2015).
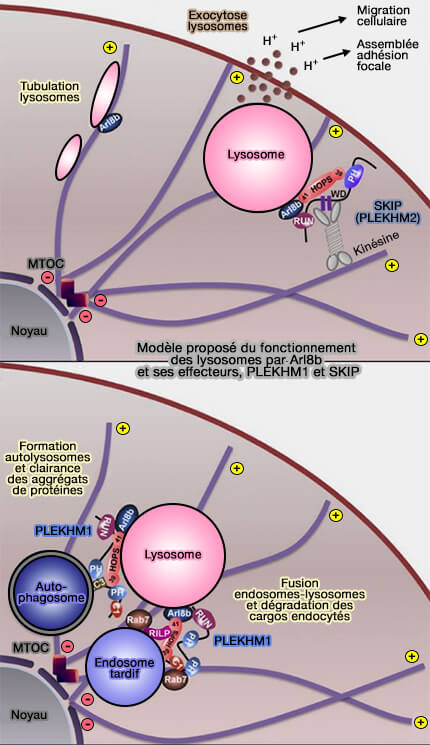
par Arl8b et PLEKHM1
(Figure : vetopsy.fr d'après Marwaha et coll)
1. PLEKHM1 interagit, comme SKIP (PLEHHM2), via leurs domaines respectifs, avec les deux GTPases Arl8b et Rab7, mais conduit les lysosomes dans un sens opposé (The Rab7 effector PLEKHM1 binds Arl8b to promote cargo traffic to lysosomes 2017).
On ne connait pas les voies de signalisation en amont et les stimuli qui conduisent à une liaison préférentielle d'un effecteur par rapport à l'autre. Toutefois, il se pourrait qu'elles dépendent de l'état nutritif de la cellule.
2. Le modèle d'activation de PLEKHM1 serait le suivant.
- a. Le complexe Rab7-RILP se lie et recrute PLEKHM1 du cytosol aux endosomes tardifs périnucléaires.
- b. PLEKHM1, via son domaine RUN, interagit avec Arl8b présent sur les lysosomes, agissant comme un lien entre les deux GTPases et repositionnant les endosomes Arl8b-positifs vers la région périnucléaire, ce qui pourrait augmenter leur accessibilité au matériel de la voie de biosynthèse (The position of lysosomes within the cell determines their luminal pH 2016).
- c. Arl8b recrute ensuite Vps41 et d'autres sous-unités (sauf Vps39) du complexe HOPS vers les sites de contact endosome/lysosome, alors que Vps39 est recruté par sa liaison directe à PLEKHM1.
Cela favorise à son tour l'attache et la fusion médiée par les protéines SNARE des vésicules des cargos avec les lysosomes ( Vps33 et SNARE).
Par exemple, la sous-unité HOPS Vps33A interagit avec la syntaxine 17 (Stx17), spécifique des autophagosomes, pour réaliser la fusion autophagosome-lysosome (The HOPS complex mediates autophagosome–lysosome fusion through interaction with syntaxin 17 2014).
Transport des endosomes/lysosomes par le cytosquelette d'actine
Biologie cellulaire et moléculaireMembrane plasmiqueNoyauCytoplasmeMitochondriesSystème endomembranaireRéticulum endoplasmiqueAppareil de GolgiEndosomes/LysosomesPeroxysomesProtéasomesCytosquelettemicrofilaments d'actineFilaments intermédiairesMicrotubulesMatrice extracellulaireReproduction cellulaireBiochimieTransport membranaireMoteurs moléculairesVoies de signalisation