


Système endo-membranaire
Réticulum endoplasmique
Sites de contact membranaire (MCS) du RE
VMP1 (vacuole membrane protein 1)
- Vue d'ensemble du système endomembranaire
- Endosomes
- Lysosomes
- Trafic des endosomes/lysosomes
- Fusion et fission membranaire
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

VMP1 (vacuole membrane protein 1) joue un rôle important dans les sites de contact membranaire (MCS) du réticulum endoplasmique.
Les progrès récents de la biologie des sites de contact membranaire (MCS) ont révélé leurs rôles clés dans les échanges interorganites.
Vue d'ensemble de VMP1
VMP1 (vacuole membrane protein 1) est une protéine spécifique des métazoaires localisée dans le réticulum endoplasmique (RE), qui joue un rôle important dans sa communication avec d’autres organites :
- omégasomes et phagophores pour la formation des autophagosomes,
- mitochondries,
- appareil de Golgi,
- gouttelettes lipidiques,
- endosomes.
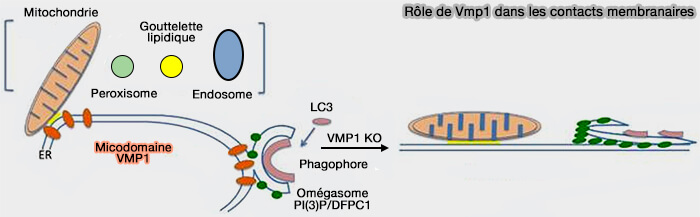
(Figure : vetopsy.fr d'après Tábara et coll)
Remarque : l’autophagie et la communication RE-membrane aux sites de contact membranaire sont étroitement liées aux troubles neurodégénératifs, tels que la maladie de Parkinson, la maladie d’Alzheimer et la sclérose latérale amyotrophique.
Structure de VMP1
1. VMP1 est une protéine qui comprend 6 domaines transmembranaires (VMP1 Establishes ER-Microdomains that Regulate Membrane Contact Sites and Autophagy 2016).
2. Les régions N- et C-terminales de VMP1 sont exposées au cytosol (The ER-Localized Transmembrane Protein EPG-3/VMP1 Regulates SERCA Activity to Control ER-Isolation Membrane Contacts for Autophagosome Formation 2017).
- La région C-terminale contient le signal de rétention RE putatif, i.e. signature xKxx.
- La région N-terminale, i.e. domaine TAG, comprend une séquence conservée avec le potentiel de former une hélice alpha amphipathique (AH) qui peut courber les membranes.
Remarque : Cette topologie est tiré des homologues KMS1 and KMS2 des plantes (KMS1 and KMS2, two plant endoplasmic reticulum proteins involved in the early secretory pathway 2011).
Rôles de VMP1
Rôles de VMP1 dans l'autophagie
VMP1 peut déclencher l'autophagie dans les cellules de mammifères (The Pancreatitis-induced Vacuole Membrane Protein 1 Triggers Autophagy in Mammalian Cells 2007).
La surexpression de VMP1 dans les cellules de mammifères induit la formation de nombreuses vésicules qui co-localisent avec LC3, un marqueur autophagosomal largement utilisé, même lors d'un apport suffisant en nutriments et en facteur de croissance (A novel mammalian trans-membrane protein reveals an alternative initiation pathway for autophagy 2008).
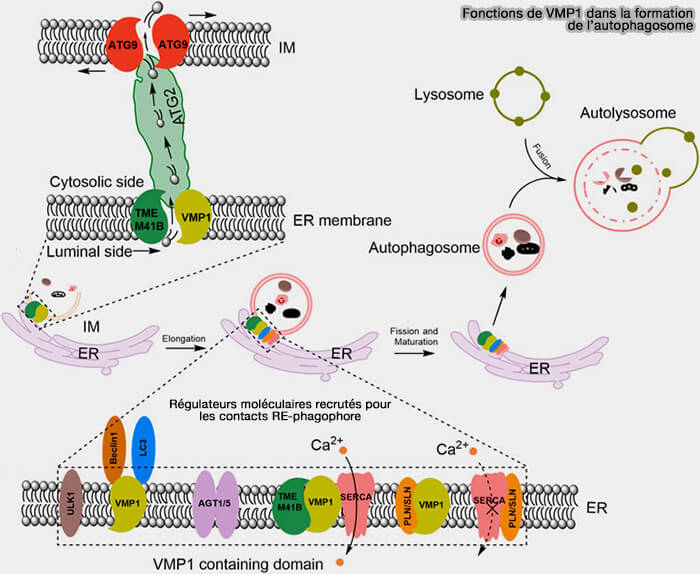
(Figure : vetopsy.fr d'après Tong et coll)
Interactions avec SERCA
1. Les réponses au stress cellulaire impliquent souvent une élévation des niveaux de calcium cytosolique, ce qui a été suggéré pour stimuler l’autophagie (The ER-Localized Transmembrane Protein EPG-3/VMP1 Regulates SERCA Activity to Control ER-Isolation Membrane Contacts for Autophagosome Formation 2017).
2. Lors de la formation d'autophagosomes dans des cellules de mammifères, les phagophores (membranes d'isolation IM) contactent dynamiquement le RE.
Dans les cellules appauvries VMP1, de nombreuses protéines ATG (AuTophaGy-related), y compris ULK1/FIP200, ATG14/Beclin1 et WIPI2, s'accumulent et se colocalisent avec LC3, et restent associées de manière stable au RE.
a. VMP1 contrôle les contacts RE-phagophore en favorisant l'activité de la pompe SERCA ou Sarco/ER Ca2+-ATPase (SERCA Control of Cell Death and Survival 2017).
- La perte de VMP1 provoque une association stable des phagophores avec le RE, bloquant ainsi la formation des autophagosomes.
- L'interaction de WIPI2 avec le complexe ULK1/FIP200 et PI(3)P contribue à la formation de contacts RE-phagophore, et ces interactions sont renforcées par la déplétion VMP1.
b. VMP1 interagit avec SERCA pour réduire les niveaux cytosoliques de Ca++ au voisinage du phagophore pour déclencher leur détachement du RE pendant la biogenèse de l'autophagosome.
- SERCA transporte le Ca++ du cytosol dans la lumière du RE pour maintenir le Ca++ cytosolique à son niveau de repos bas.
- VMP1 inhibe la formation du complexe inhibiteur de SERCA/PLN/SLN, i.e. phospholamban (PLN) et sarcolipine (SLN), protéines membranaires intégrales du RE,qui interagissent avec SERCA pour réduire son affinité pour le Ca++ cytosolique.
c. Le traitement par la thapsigarine (TG), un inhibiteur spécifique du SERCA, arrête la biogenèse de l'autophagosome avant le détachement de phagophore/autophagosomes du RE (Modulation of intracellular calcium homeostasis blocks autophagosome formation 2013).
- Une faible concentration de T,, qui ne provoque pas de changements durables ou détectables dans les niveaux de Ca++ cytosoliques, bloque toujours l'autophagie, suggérant que le mécanisme inhibiteur pourrait impliquer des perturbations plus subtiles ou locales de Ca++ à proximité du phagophore sur le RE.
- La façon dont le taux de Ca++ est régulé pendant la dissociation reste inconnue.
3. La modification des niveaux de Ca++ pourrait moduler l'activité VPS34 du complexe PI3KC3-CI au niveau des sites de formation des autophagosomes.
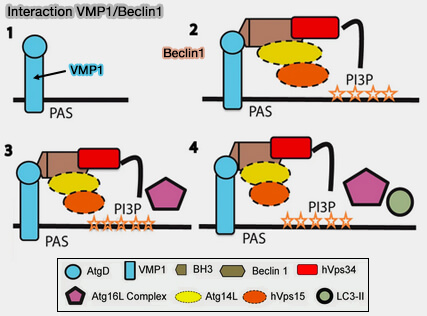
(Figure : vetopsy.fr d'après Molejon et coll)
a. VMP1 interagit avec le domaine BH3 de Beclin1 du complexe et la région C-terminale hydrophile de la VMP1 (domaine TAG) est essentielle pour son interaction avec Beclin-1 et l'induction de l'autophagie (The VMP1-Beclin 1 interaction regulates autophagy induction 2013).
L'expression de VMP1 avec un domaine ATG muté ne parvient pas à induire le recrutement et l'interaction de la LC3 avec Beclin-1, qui abolit les triples co-localisations de VMP1, LC3 et Beclin-1.
b. Cela pourrait diminuer le niveau de PI(3)P sur les sites de formation d'autophagosomes, contribuant à la dissociation du complexe de fixation RE-phagophore médiée par WIPI2.
Remarque : VMP1 interagirait aussi avec ATG2 pour le transfert des lipides au phagophore.
Interactions avec ATG2 pour le transfert des lipides
1. L’autophagosome pourrait se développer même à partir d’une seule vésicule contenant ATG9, agissant comme une membrane " graine " (A model for a partnership of lipid transfer proteins and scramblases in membrane expansion and organelle biogenesis 2021).
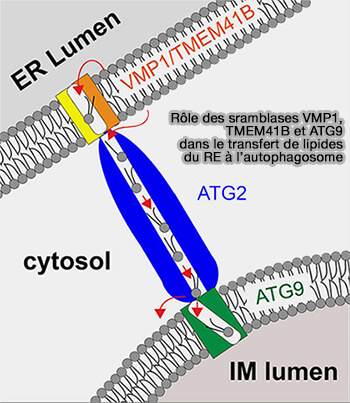
Figure: vetopsy.fr d'après Ghanbarpour et coll)
a. ATG2 permettrait le transport des lipides du réticulum endoplasmique (RE) vers la vésicule d’ensemencement.
- TMEM41B et VMP1, qui sont deux scramblases, rééquilibreraient les feuillets du RE lors de l’extraction des lipides (
 cf plus bas).
cf plus bas). - ATG9, autre scramblase, équilibrerait les lipides dérivés du RE dans la " graine " au fur et à mesure qu’ils sont délivrés (Structure, lipid scrambling activity and role in autophagosome formation of ATG9A 2020).
b. Cette redistribution permettrait d’élargir la surface de la membrane de la graine, même si le volume du contenu enfermé dans la membrane reste relativement constant.
Rôles de VMP1 dans les MCS

Vous pouvez lire : Roles of VMP1 in Autophagy and ER–Membrane Contact: Potential Implications in Neurodegenerative Disorders (2020) et VMP1: a multifaceted regulator of cellular homeostasis with implications in disease pathology (2024).
VMP1, mais aussi TMEM41B, ont tous deux été impliqués dans de multiples processus autres que l’autophagie, tous potentiellement associés d’une manière ou d’une autre à la dynamique membranaire.
1. Elles sont impliquées dans l’homéostasie lipidique, car la déplétion de l’un ou l’autre conduit à l’accumulation de lipides neutres dans des gouttelettes lipidiques cytoplasmiques surdimensionnées (TMEM41B is a novel regulator of autophagy and lipid mobilization 2018).
2. Les deux protéines sont enrichies aux sites de contact organite-organite avec la machinerie clé de synthèse des lipides.
Elles sont donc idéalement situées pour faciliter la redistribution des lipides du RE via des protéines solubles de transfert de lipides localisées aux sites de contact (Vacuole membrane protein 1 marks endoplasmic reticulum subdomains enriched in phospholipid synthesizing enzymes and is required for phosphoinositide distribution 2018).
3. VMP1 est essentiel à la production de lipoprotéines (A critical role of VMP1 in lipoprotein secretion 2019).
a. L’assemblage des lipoprotéines a lieu dans la lumière du RE, où les apolipoprotéines sont assemblées avec des lipides dérivés de manière asymétrique du feuillet luminal de la membrane du RE avant d’être emballées dans des vésicules pour être transportées hors de la cellule.
L’appauvrissement de VMP1 conduit à la production de particules amorphes de type lipoprotéine qui suggèrent un bourgeonnement partiel dans les feuillets cytoplasmique et luminal simultanément, et une perte complète de sécrétion efficace.
b. Cela suggère que, de manière analogue au couplage des scramblases et des protéines de transport des lipides cytoplasmiques, il existe un couplage fonctionnel comparable de l’activité des scramblases et de la construction efficace des particules de lipoprotéines.
Remarque : les coronavirus et les flavivirus forment des compartiments de réplication dérivés de la membrane du RE, décrits comme des structures sphériques à double membrane, ressemblent à des autophagosomes et pourraient se former de novo, comme l’autophagosome, impliquant un partenariat entre les protéines de transfert de lipides et les scramblases (TMEM41B Is a Pan-flavivirus Host Factor 2021).
Retour au réticulum endoplasmique ou à l'autophagie
Biologie cellulaire et moléculaireMembrane plasmiqueNoyauCytoplasmeMitochondriesSystème endomembranaireRéticulum endoplasmiqueAppareil de GolgiEndosomesLysosomesPeroxysomesProtéasomesCytosquelettemicrofilaments d'actineFilaments intermédiairesMicrotubulesMatrice extracellulaireReproduction cellulaireBiochimieTransport membranaireMoteurs moléculairesVoies de signalisation