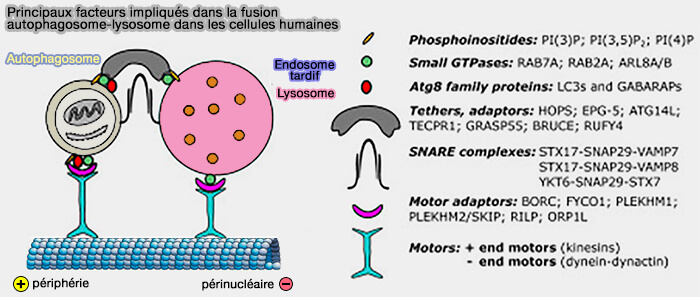
La fusion des autophagosomes avec les différents compartiments, endosomes tardifs et lysosomes, nécessite une machinerie de fusion conduite par les protéines SNARE.
Principaux facteurs impliqués dans la fusion des autophagosomes
(Figure : vetopsy.fr d'après Lőrincz et Guhász)
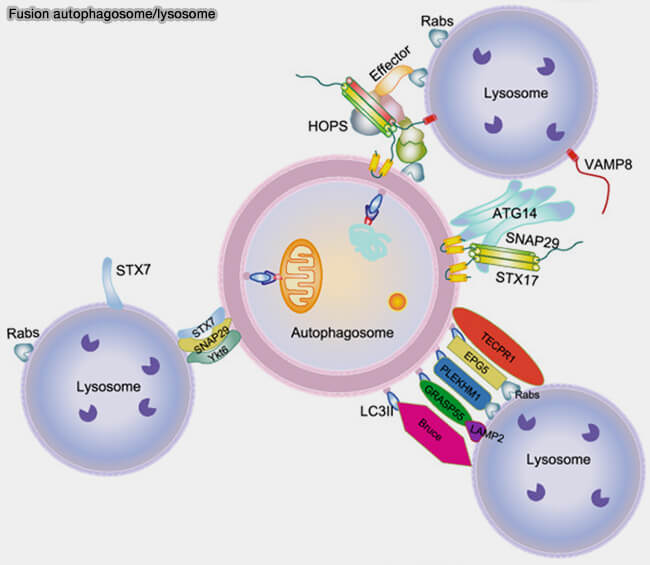
Fusion autophagosome/lysosome
Fusion autophagosome/lysosome
(Figure : vetopsy.fr d'après Li et coll)
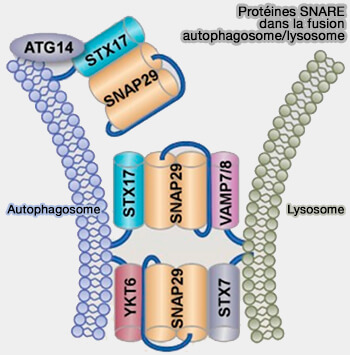
La protéine d'autophagie associée à l'autophagosome ATG14 interagit avec STX17 pour favoriser l'assemblage et la stabilisation du sous-complexe STX17/SNAP29 pour le préparer à l'interaction avec VAMP8 ( rôle d'ATG14 dans la fusion autophagosome/lysosome).
a. Le premier modèle est que YKT6 et STX17 se localisent dans les autophagosomes indépendamment l’un de l’autre et forment chacune un complexe SNARE différent pour la fusion autophagosome/lysosome.
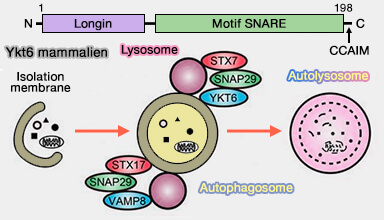
Ykt6 mammalien
(Figure : vetopsy.fr d'après Baker et coll)
YKT6 forme un complexe SNARE avec STX7-SNAP29.
STX17 forme un complexe SNARE avec SNAP29-VAMP8.
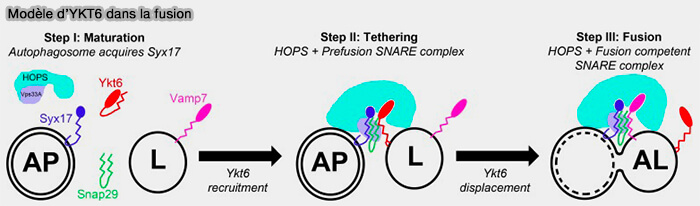
b. Le deuxième modèle pourrait être le suivant.
1. Au cours de l’étape de maturation, les autophagosomes naissants acquièrent une compétence de fusion en recrutant Syx17.
2. Au stade d'attache, Ykt6 soluble est recruté et s’associe à la membrane lysosomale.
De manière concomitante, Ykt6 peut former un complexe SNARE incompétent et pas complètement zippé avec Syx17 et Snap29, qui peut fournir de multiples surfaces pour l’interaction et le recrutement du complexe HOPS.
3. Lors de l'étape de la fusion, Vamp7 déplace Ykt6 et forme un complexe SNARE stable et compétent avec Syx17 et Snap29 pour réaliser la fusion membranaire.
Modèle d'action d'EPG5
(Figure : vetopsy.fr d'après Wang et coll)
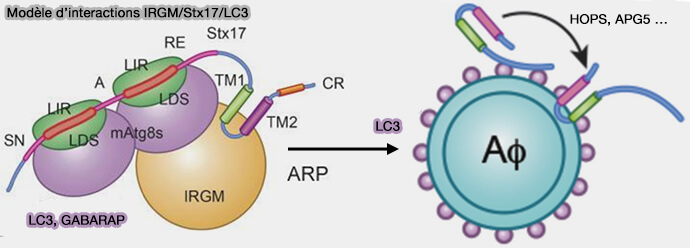
Modèle d’interactions IRGM/Stx17/LC3
(Figure : vetopsy.fr d'après Kumar et coll)
2. Un modèle simplifié de la façon dont la particule de reconnaissance des autophagosomes (ARP) fournit Stx17 aux autophagosomes pourrait être le suivant.
Tout d’abord, l’ARP, i.e. IRGM, Stx17 avec leurs domaines LIR liés à LC3 ou à d’autres Atg8 et peut-être à plus d’autres composants potentiels, permet l’insertion de Stx17 dans la membrane autophagosomale par des interactions d’échange avec LC3/Atg8 sur les autophagosomes.
Ensuite, le domaine SNARE, occupé par LC3/Atg8, est libéré de manière contrôlée, par exemple, par le complexe HOPS avec d’autres facteurs clés tels que EPG5…, pour former un complexe SNARE avec d’autres SNARE.
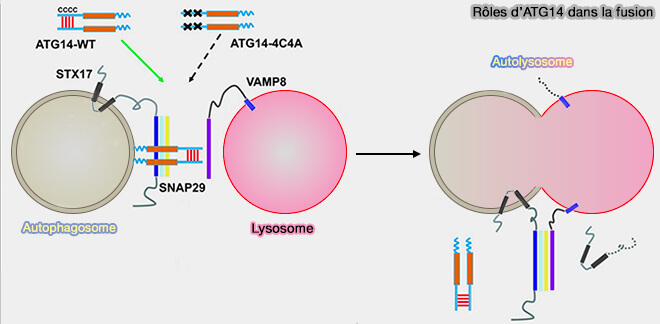
Rôle d'ATG14 dans la fusion
(Figure : vetopsy.fr d'après Liu et coll)
L’ATG14 est homo-oligomérisé par ses répétitions de cystéine.
L’oligomère ATG14 cible les autophagosomes complets et interagit avec une hélice α du domaine central SNARE de STX17 par le biais de son domaine coiled-coil.
Il stabilise le complexe binaire STX17-SNAP29, le préparant à se lier à VAMP8 sur les lysosomes pour favoriser la fusion entre les autophagosomes et les lysosomes.
Après fusion, ATG14 se dissocie du complexe cis-SNARE et est récupéré à partir des autolysosomes.
Dans les cellules ATG14 défectueuses d’homo-oligomérisation, les autophagosomes se forment toujours efficacement, mais leur fusion avec les endolysosomes est bloquée.
STX17 est situé au niveau de la membrane externe des autophagosomes complets grâce à une structure en épingle à cheveux C-terminale.
Elle n'est pas présente sur les phagophores, i.e. le lysosome ne peut pas fusionner avec les premières structures autophagiques.
2. Deux minutes seulement après le recrutement de STX17, les petits lysosomes commencent à fusionner avec les autophagosomes, suggérant une acidification de l'espace entre la membrane autophagosomale interne et la membrane autophagosomale externe.
Environ sept minutes plus tard, la matrice autophagosomale a été complètement acidifiée, ce qui suggère que la membrane autophagosomale interne est dégradée.
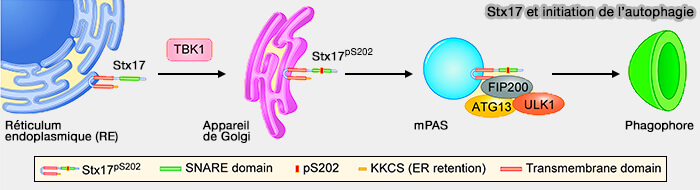
Lors de l'induction de l'autophagie, Stx17pS202 se déplace de l'appareil de Golgi au mPAS, i.e. structure pré-autophagosomale, et contrôle la formation des structures PAS positives à FIP200 et ATG13.
Stx17pS202 serait nécessaire pour les interactions robustes entre les composants du complexe FIP200-ATG13-ULK1.
Stx17 et initiation de l'autophagie
(Figure : vetopsy.fr d'après Kumar et coll)
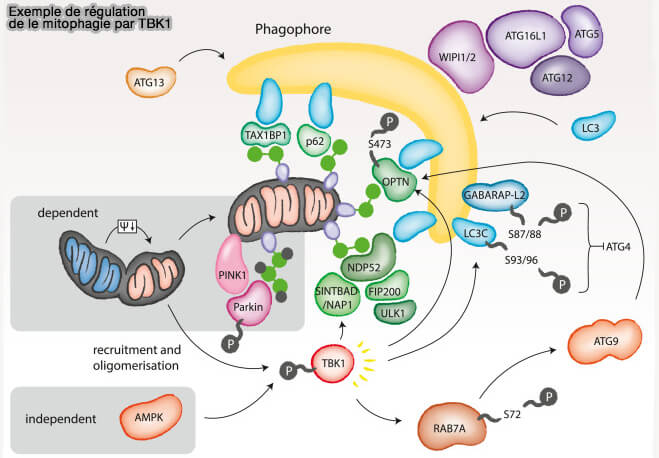
Étant donné que le KO de TBK1 a réduit cette interaction, il est probable que la phosphorylation par TBK1 de Stx17 affecte la capacité de Stx17 à s'associer aux premières structures autophagosomales, contribuant éventuellement aux transitions entre les autophagosomes mPAS et LC3-positifs.
À l'appui de ce point de vue, Stx17pS202 se co-localise également avec DFCP1, un marqueur pour une structure intermédiaire d'omégasomes d'où émergent des autophasomes naissants.
Autophagie et TBK1
(Figure : vetopsy.fr d'après Kumar et coll)
et après la fusion ?
L'autolysosome, par son acidité et ses hydrolases lysosomales, pourra digérer les différents constituants.
La macroautophagie génère des nucléotides, des acides aminés et des acides gras libres qui pourront être réutilisés pour la synthèse de nouvelles macromolécules et d’énergie (ATP).
Dérégulations de la maturation et la fusion des autophagosomes
Les dérégulations de la maturation et la fusion des autophagosomes sont impliquées dans :
les maladies neurodégénératives,
les maladies musculaires,
les cancers,
les maladies par agents pathogènes qui utilisent diverses stratégies pour inhiber l'autophagie aux étapes d'initiation et/ u de maturation pour échapper à la destruction.
Certains agents pathogènes bloquent même la maturation des autophagosomes et renversent les vésicules résultantes pour leur propre avantage.





 rôle d'ATG14 dans la fusion autophagosome/lysosome).
rôle d'ATG14 dans la fusion autophagosome/lysosome).