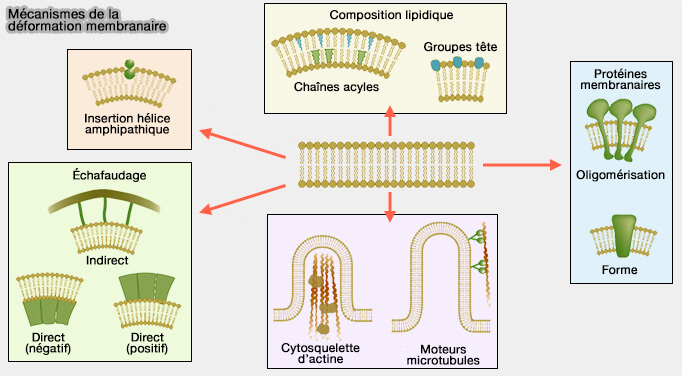
Mécanisme de déformation membranaire
(Figure : vetopsy.fr d'après Mc Mahon et coll)
Les hélices amphipathiques (AH), caractérisées par la présence de résidus basiques à l'interface polaire-non polaire de leurs deux faces, sont bien adaptées à l'interaction avec les membranes.
Les AH enfoncent un " coin " dans la membrane (wedging mechanism).
Remarque : les hélices hydrophobes diffèrent des AH car tous leurs résidus sont hydrophobes.
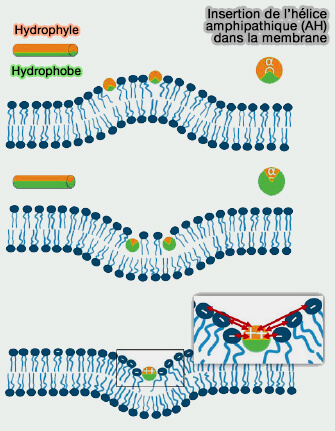
Insertion des hélices amphipathiques (AH)
(Figure : vetopsy.fr d'après Jin et coll)
un côté est polaire (chargé),
l'autre hydrophobe.
2. Lors de l'insertion membranaire, la face hydrophobe de l'AH est enterré à l'intérieur de la membrane hydrophobe et la face polaire est exposée aux groupes de tête lipidiques hydrophiles et à la phase aqueuse.
Ce phénomène provoque le déplacement des groupes de tête lipidiques et une réorientation des chaînes acyles, donnant une orientation plus favorable à une courbure plus élevée.
L'AH insérée dans la membrane peut faciliter la génération de structures membranaires courbes, y compris des vésicules, des tubules et même des membranes en forme de selle.
Les résidus chargés sur la face polaire complique les interactions AH-membrane et contribue en outre à la diversité des phénomènes de remodelage membranaire induits par les AH.
3. En outre, les AH de nombreuses protéines induisant la fission contiennent :
au moins deux résidus basiques, i.e. arginine (R) et/ou lysine (K) chacun, à l'interface polaire-non polaire, i.e. Lys7 et Lys16 pour l'endophiline…
au moins trois résidus chargés positivement.
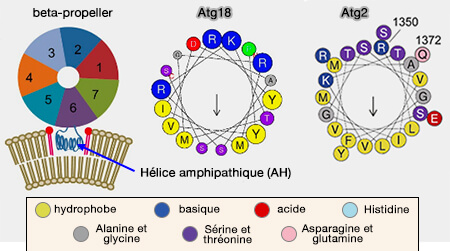
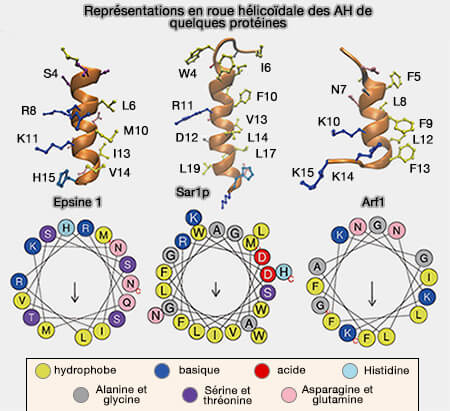
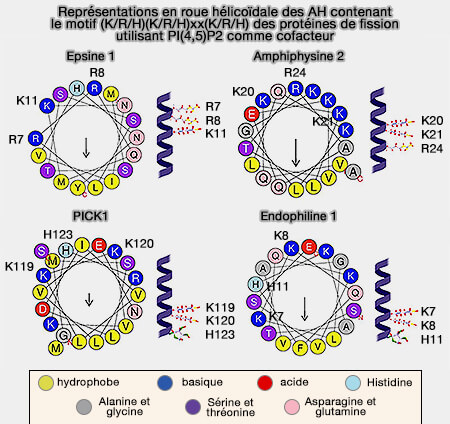
Hélices amphipathiques (AH) de quelques protéines
(Figure : vetopsy.fr d'après Zhen-lu et coll)
Les AH des protéines qui détectent, mais ne génèrent pas de courbure membranaire, comme ArfGAP1… ne contiennent souvent pas plus d'un résidu basique.
L'interaction des AH des protéines induisant la fission avec des lipides spécifiques pourrait assurer la profondeur d'insertion optimale pour la génération de la courbure de la membrane et pour l'interaction avec d'autres protéines induisant la fission.
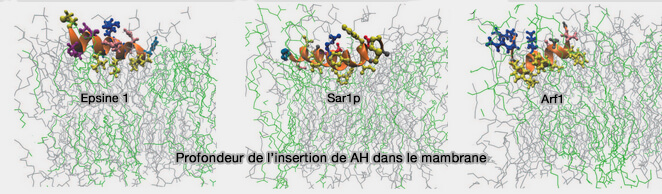
Profondeur de l'insertion des hélices amphipatiques (AH) dans la membrane
(Figure : vetopsy.fr d'après Mc Mahon et coll)
2. Un mécanisme supplémentaire prévoit une coopérativité entre les protéines où la première insertion d'un AH à partir d'une protéine donnée capable de détecter la courbure de la membrane, i.e. de se lier sélectivement aux membranes courbes, favorise l'insertion d'AH supplémentaires (Sar1 GTPase Activity Is Regulated by Membrane Curvature 2016).
À leur tour, ces AH induisent plus de courbure membranaire facilitant l'insertion d'AH supplémentaires.
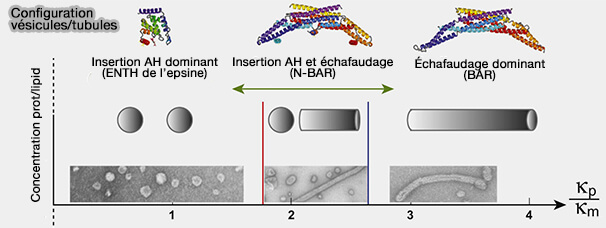
Wedging versus Scaffolding mecanims
(Figure : vetopsy.fr d'après Boucrot et coll)
En raison de cette coopérativité positive dans la génération de courbure membranaire, le processus d'emballement conduisant à la fission membranaire pourrait avoir lieu une fois qu'une concentration critique de protéines induisant la fission contenant AH est atteinte.
Remarque : les mécanismes de formation des vésicules ou tubules des protéines BAR sont étudiés avec les fonctions des domaines BAR.
Cofacteurs lipidiques d'insertion
Hélice amphipathique (AH) et PI(4,5)P2
(Figure : vetopsy.fr d'après Zhukovsky et coll)
1. De nombreuses protéines à AH induisant la fission nécessitent un ou deux lipides spécifiques pour générer une courbure membranaire et terminer le processus de fission.
La plupart de ces lipides sont anioniques.
Ces lipides pourraient être nommés cofacteurs lipidiques, facteurs lipidiques ou ligands lipidiques.
Certaines petites GTPases possèdent un hélice amphipathique (AH) N-terminale, qui lors de l'échange GDP/GTP, subissent un changement conformationnel qui expose l'AH pour son insertion membranaire.
Hélices amphipathiques (AH) de quelques protéines
(Figure : vetopsy.fr d'après Zhen-lu et coll)
Sar1-GTP activé par Sec12 s'insère dans la membrane du RE, se liant préférentiellement aux zones de courbure de la membrane.
Lorsque Sar1-GTP s'insère dans la membrane, il recrute Sec23 et Sec24 pour constituer la cage interne de COPII.
Protéines à domaine ENTH/ANTH
1. L'epsine se lie par son domaine ENTH au PI(4,5)P2 ou PIP2 membranaire, ce qui provoque un changement de conformation qui ajoute une hélice α0 ou H0 à la fin du domaine.
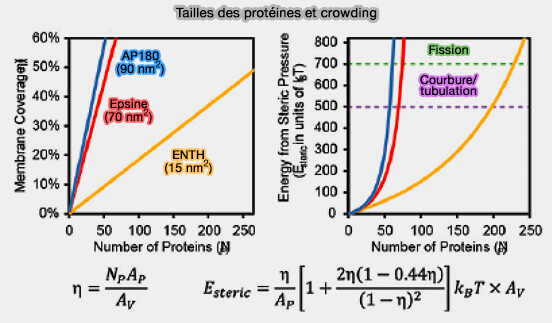
La capacité du domaine ENTH à former des tubules membranaires et la vésiculation de la membrane pour forment des vésicules plus petites serait corrélée à la couverture de la surface membranaire par les protéines plutôt qu'à l'insertion d'hélices amphipathiques dans les membranes ( crowding et courbure/fission).
Taille des protéines et crowding
(Figure : vetopsy.fr d'après Zeno et et Stachowiak)
2. Atg18 forme un complexe avec Atg2 et joue également un rôle dans la formation des autophagosomes et intervient également dans la fission des vacuoles.
Remarque : contrairement aux petites GTPasesArf1 et Sar1, aux protéines à domaine ENTH/ANTH, aux protéines N-BAR contenant toutes des AH N-terminaux, les AH des des protéines Atg2 et Atg18, similaires à l'AH de PICK1, sont internes et loin de l'extrémité N-terminale.
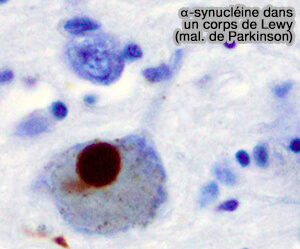
Synucléines
Les synucléines sont une famille de protéines solubles communes aux vertébrés, principalement exprimées dans le tissu neuronal, mais aussi dans certaines tumeurs.
α-synucléine dans un corps de Lewy
(Figure : vetopsy.fr d'après Marvin 101)






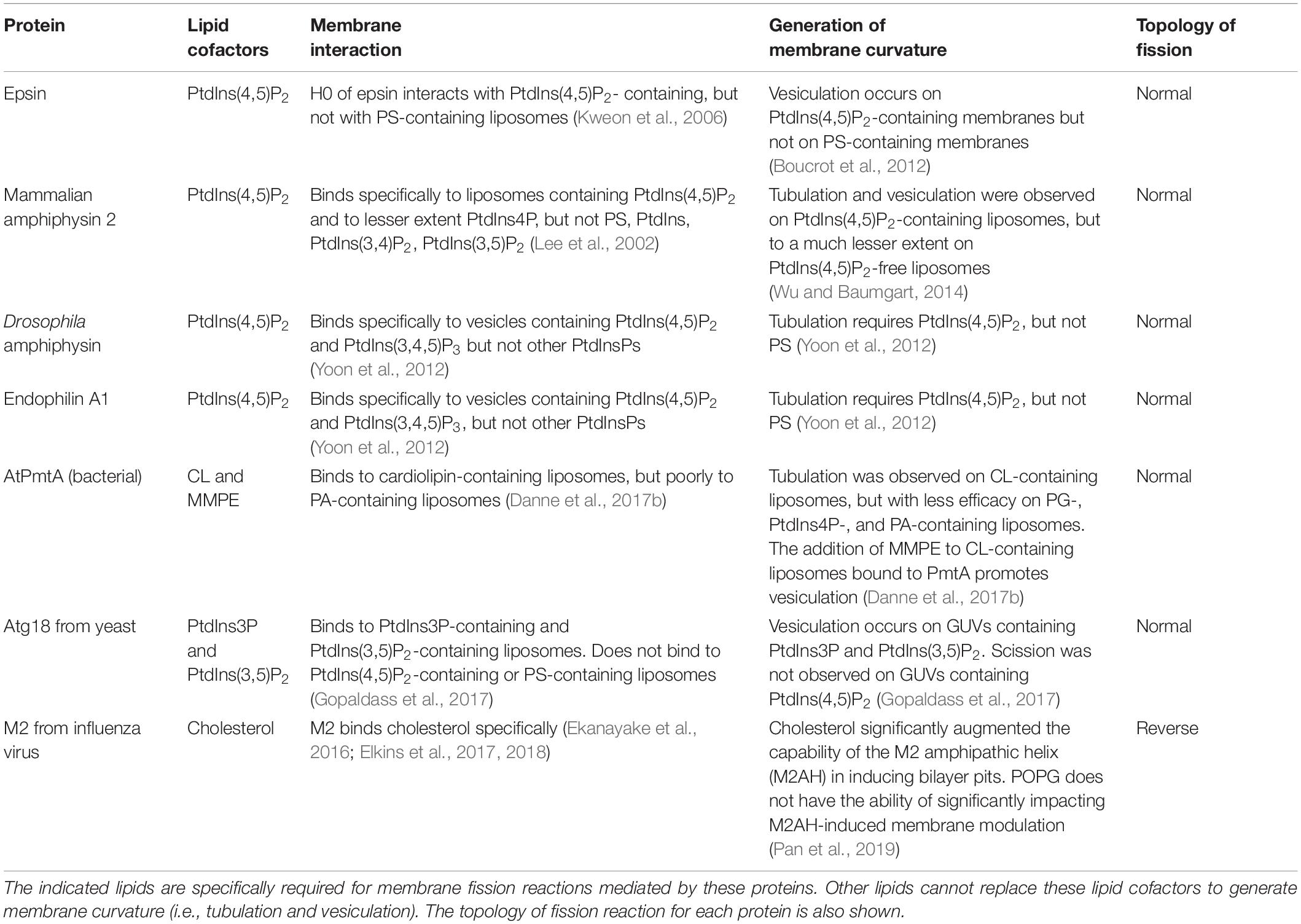 cf. tableau) :
cf. tableau) :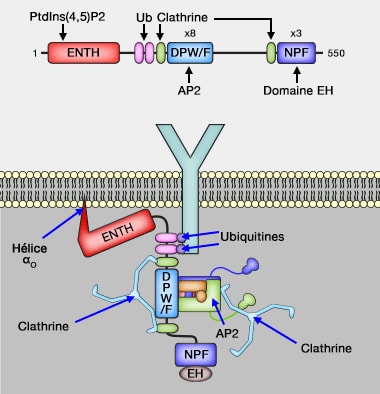
 crowding et courbure/fission).
crowding et courbure/fission).