


Autophagie
Protéines ATG (AuTophaGy-related)
Atg2
- Vue d'ensemble du système endomembranaire
- Autophagie
- Système UPS (Système UPS (Ubiquitine-protéasome))
- Fusion et fission membranaire
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

ATG2 est une protéine de transfert des lipides (LTP ou Lipid Tranfer Protein) impliquée dans l'élongation du phagophore.
Les LTP, protéines de transfert des lipides (LTP ou Lipid Tranfer Protein) intracellulaires se localisent sur les sites de contact membranaire (MCS) au sein desquels les LTP peuvent transférer les lipides d'un compartiment donneur à un compartiment accepteur.
Vue d'ensemble d'ATG2
ATG2, comme Vps13, est une LTP de très grande taille, 1 500 à 3 000 résidus, par rapport aux LTP tubulaires (TULIP).
ATG2 appartient à la famille des protéines à domaine choréine, appelée ainsi car le VPS13A humain est aussi appelé choréine.
1. ATG2 peut relier deux membranes par un long pont protéique en forme de tige avec une rainure hydrophobe à travers laquelle les lipides peuvent glisser alors que la protéine reste stationnaire.
a. Ce mécanisme est parfaitement adapté au transport en vrac des lipides bicouches pour favoriser la croissance membranaire.
b. Un modèle prédit un transfert de 100 lipides par seconde (Kinetics of Atg2-mediated lipid transfer from the ER can account for phagophore expansion 2020).
La génération d'autophagosomes avec un diamètre de ~ 400 nm nécessite environ 3 millions de lipides, ce qui nécessiterait ~ 90 molécules ATG2 pour fournir suffisamment de lipides en 10 min.

(Figure : vetopsy.fr d'après plusieurs sources)
2. ATG2, impliquée dans le site de l'assemblage du phagophore (PAS) dans l'autophagie, à structure plus courte que VPS13 (d'environ 1500 à 2300 résidus), a une conformation allongée comportant un canal (ATG2 transports lipids to promote autophagosome biogenesis 2019).
Atg2 de la levure a deux homologues chez l'homme (ATG2A/B).
Structure d'ATG2
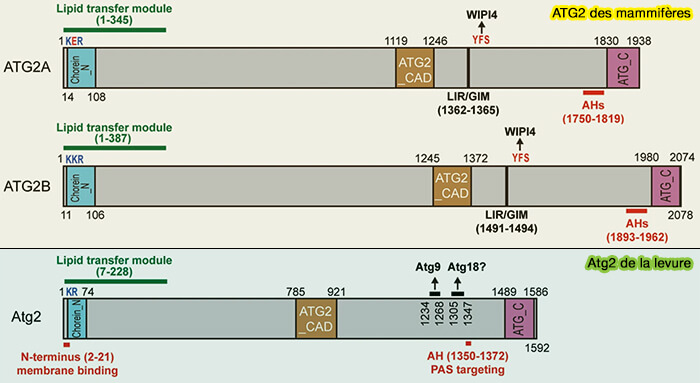
(Figure : vetopsy.fr d'après Melia et coll)
1. Sa extrémité N-terminale comprend un domaine choréine très proche homologiquement de celui de VPS13 impliqué dans les sites de contact membranaires (MCS) entre organites.
- Cette région de liaison de la membrane N-terminale est essentielle pour la formation d'autophagosomes (Differential requirement for ATG2A domains for localization to autophagic membranes and lipid droplets 2017).
- Elle permet le transport des lipides, en particulier la phosphatidyléthanolamine (PE) du réticulum endoplasmique (RE)/omégasome jusqu'au phagophore en croissance (ATG2 transports lipids to promote autophagosome biogenesis 2019).
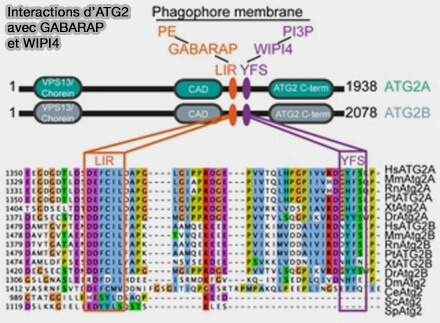
2. ATG2 contient un motif CAD (Cys-Ala-Asp ou YFS), unique à ATG2 et étroitement lié à son extrémité à WIPI4, dont le rôle est encore obscur ( interactions ATG2/WIPI4).
interactions ATG2/WIPI4).

(Figure : vetopsy.fr d'après Maeda et coll)
3. Suit un motif LIR (LC3 Interacting Region) qui pourrait se lier à ATG18, i.e. LIR5 d'ATG2A et LIR6 d'ATG2B ( tableau).

(Figure : vetopsy.fr d'après s Kolokowski et coll)
Certains auteurs prédisent la présence de plusieurs domaines LIR, i.e. 5 dans ATG2A et 6 dans ATG2B (A conserved ATG2‐GABARAP family interaction is critical for phagophore formation 2020), mais un seul serait lié à cette interaction ( problématique des domaines LIR).
4. Le domaine APT1 se lie aux phosphoinositides, i.e. PI(3)P (The binding of the APT1 domains to phosphoinositides is regulated by metal ions in vitro 2020).
Chez VPS13, il fait partie du domaine DHL/PH.
4. Son extrémité C-terminale présente deux séquences remarquables.
a. un court segment de similarité de séquence primaire avec celle de Vps13, i.e. ATG2_C (1724-1829) chez ATG2A, qui comprend une hélice amphipathique i.e. 1750-1767 permettant la localisation aux différents organites dont les autophagosomes (Differential requirement for ATG2A domains for localization to autophagic membranes and lipid droplets 2017).
-
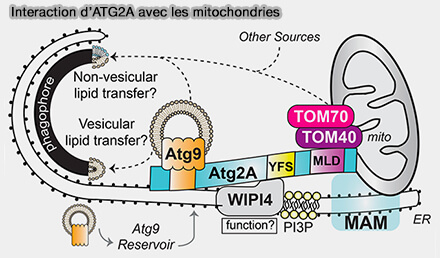
Interaction ATG2A avec TOM40 des mitochondries
(Figure : vetopsy.fr d'après Tang et coll) - Cette dernière implique la liaison à TOM40, une protéine de la membrane mitochondriale externe (TOM40 Targets Atg2 to Mitochondria-Associated ER Membranes for Phagophore Expansion 2019).
b. Un autre segment (1830-1938) serait impliqué dans la localisation aux gouttelettes lipidiques (LD), mais pas pour l’autophagie.
Partenaires pour la liaison membranaire d'ATG2
ATG2 ne possède pas de domaine WD40 pour se lier à la membrane et le recrutement spécifique d'Atg2/ATG2B au phagophore nécessite des partenaires.
1. À son extrémité N-terminale, elle contacte (A model for a partnership of lipid transfer proteins and scramblases in membrane expansion and organelle biogenesis 2021) :
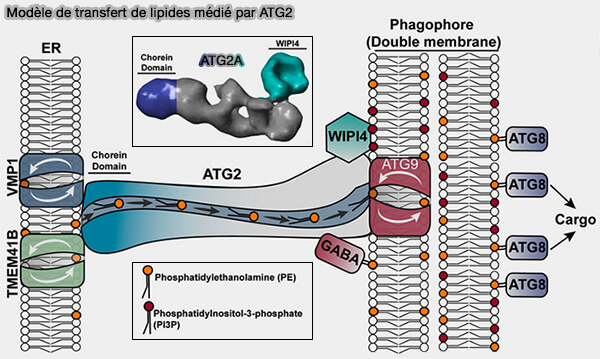
(Figure : vetopsy.fr d'après MacEwan et coll)
2. À son extrémité C-terminale, elle interagit avec plusieurs protéines.
a. Atg9 forme un complexe hétérotétramérique, i.e. domaines C-terminal du trimère d'Atg9 et domaine N-terminal d'Atg2 ( liaison ATG2-ATG9).
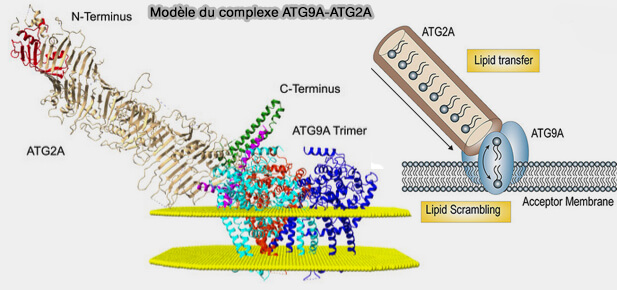
(Figure : vetopsy.fr d'après Van Vliet et coll)
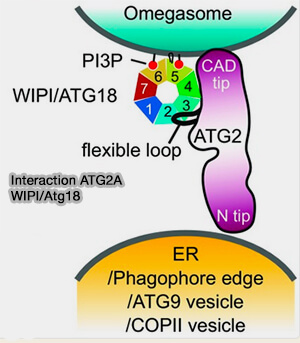
b. WIPI4/Atg18 se lie au domaine CAD d'ATG2 ( interactions ATG2/WIPI4).
Elle permet de cibler ATG2 sur les membranes enrichies en PI(3)P du phagophore grâce à son hélice amphipathique (AH) et ses deux sites de liaison ( liaisons membranaires de WIPI).
(Figure : vetopsy.fr d'après Bozic et coll)
c. GABARAP se lie directement par son domaine LIR à ATG2A et ATG2B (A conserved ATG2‐GABARAP family interaction is critical for phagophore formation 2020).
- Cette séquence LIR inhabituelle, i.e. FCIL, est située à une trentaine de résidus du motif CAD (Cys-Ala-Asp ou YFS) de la liaison à WIPI4.
- Les interactions entre ATG2, GABARAP et WIPI4 sont indépendantes.
d. TRAPP-III, et en particulier TRAPPC11 semble réguler le recrutement d'ATG2B et de WIPI4 au phagophore et implique son extrémité C-terminale.
- La déplétion de TRAPC11 se traduit par un phénotype similaire à celui des KO de l'ATG2A/B et de l'ATG2A-mLIR.
Remarque : ATG2 et TRAP-III sont nécessaires également à la fermeture de l'autophagosome.
(Figure : vetopsy.fr
d'après Chowdhury et coll)
3. En outre, ATG2A interagit avec TOM40/TOM70 sur les mitochondries ( cf. plus haut).
Rôles d'ATG2
ATG2 a plusieurs rôles dans l'autophagie.
Transfert de lipides
ATG2 permet le transfert des lipides dans l'autophagosome en formation.
1. L’autophagosome pourrait se développer même à partir d’une seule vésicule contenant ATG9, agissant comme une membrane " graine " (A model for a partnership of lipid transfer proteins and scramblases in membrane expansion and organelle biogenesis 2021).
a. ATG2 permettrait le transport des lipides du réticulum endoplasmique (RE) vers la vésicule d’ensemencement.
- TMEM41B et VMP1, qui sont deux scramblases, rééquilibreraient les feuillets du RE lors de l’extraction des lipides.
-
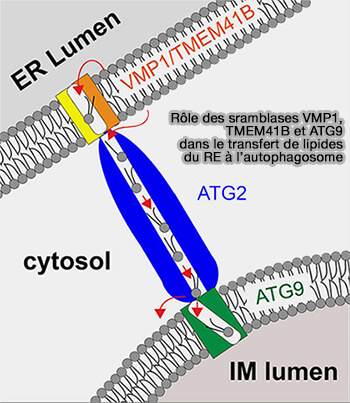
Rôles des scramblases dans le transfert lipidique
Figure : vetopsy.fr d'après Ghanbarpour et coll)
b. Cette redistribution permettrait d’élargir la surface de la membrane de la graine, même si le volume du contenu enfermé dans la membrane reste relativement constant.
2. Une telle expansion entraînerait une structure à double membrane comme l’autophagosome.
La courbure membranaire élevée est coûteuse en énergie, de sorte que l’autophagosome naissant ne formerait pas une double membrane, avec une circonférence expansive à forte courbure, mais s’enroulerait spontanément en une structure en forme de coupe, avec une zone de courbure élevée beaucoup plus petite, comme observé dans l’autophagosome en maturation.
Remarque : VMP1 et TMEM41B ont tous deux été impliqués dans de multiples processus autres que l’autophagie, tous potentiellement associés d’une manière ou d’une autre à la dynamique membranaire ( rôles de VMP1 et de TMEM41B dans les processus lipidiques).
Fermeture de l'autophagosome

Les mécanismes de la fermeture de l'autophagosome sont mal compris.
1. La déplétion simultanée d'ATG2A et d'ATG2B entraîne l'accumulation de petites structures de phagophores immatures ouvertes (Mammalian Atg2 proteins are essential for autophagosome formation and important for regulation of size and distribution of lipid droplets 2012).
a. La déplétion du WIPI4 provoque également des structures ouvertes de phagophore, mais elles sont morphologiquement différentes de celles générées après la déplétion d'ATG2A/B2 (WIPI3 and WIPI4 β-propellers are scaffolds for LKB1-AMPK-TSC signalling circuits in the control of autophagy 2017).
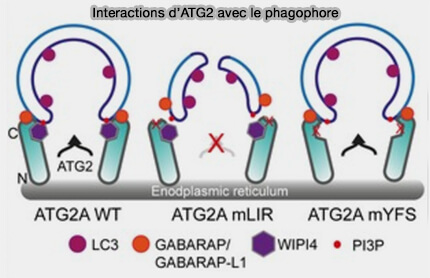
(Figure : vetopsy.fr d'après Bozic et coll)
On ne sait pas si l'interaction ATG2-WIPI4 est nécessaire pour la restauration du flux d'autophagie dans les cellules déprivées en ATG2A/B.
b. La perturbation de l'interaction ATG2A-WIPI4 n'a pas eu d'effets perceptibles sur la fermeture du phagophore et le flux d'autophagie.
2. La mutation du motif LIR sur ATG2 a complètement bloqué la fermeture du phagophore et le flux d'autophagie, malgré la possibilité de l'interaction ATG2/WIPI4 (A conserved ATG2‐GABARAP family interaction is critical for phagophore formation 2020).
Étant donné que le mutant ATG2-LIR a altéré le flux d'autophagie, mais peut encore se localiser aux gouttelettes lipidiques (LD), il se pourrait que l'ATG2-AH et l'ATG2-LIR soient essentiels pour définir la membrane cible, permettant le transfert de lipides et la formation efficace de phagophore et la maturation des autophagosomes.
Retour à l'expansion membranaire du phagophore
Biologie cellulaire et moléculaireConstituants de la celluleSystème endomembranaireRéticulum endoplasmiqueAppareil de GolgiEndosomesLysosomesPeroxysomesAutophagieMacroautophagieMicroautophagieAutophagie chaperonnes (CMA)Autophagie non canoniqueProtéines ATGSystème UPS (Ubiquitine-Protéasome)Transport membranaireTrafic vésiculaireFusion/fission membranaireEndocytoseVoie sécrétoireMoteurs moléculairesVoies de signalisation