


Autophagie
Autophagie non canonique
CASM (Conjugation of ATG8 to Single Membranes)
Vue d'ensemble
- Vue d'ensemble du système endomembranaire
- Autophagie
- Système UPS (Système UPS (Ubiquitine-protéasome))
- Fusion et fission membranaire
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

L'autophagie non canonique par la lipidation Atg8/LC3-II est différente de l'autophagie canonique et ATG8/LC3-II se lie à des membranes simples et non doubles.
Autophagie non canonique versus autophagie canonique
1. L'autophagie non canonique par la lipidation Atg8/LC3-II est différente de l'autophagie canonique.

Dans cette autophagie, ATG8/LC3-II se lie à des membranes simples et non doubles comme dans l'autophagosome, pour marquer les compartiments intracellulaires endommagés afin de signaler le danger causé par des agents pathogènes ou des composés toxiques.
2. L'autophagie non canonique par Atg8/LC3-II peut afficher divers profils.
a. La plus fréquente est appelée CASM, i.e. Conjugation of ATG8 to Single Membranes.
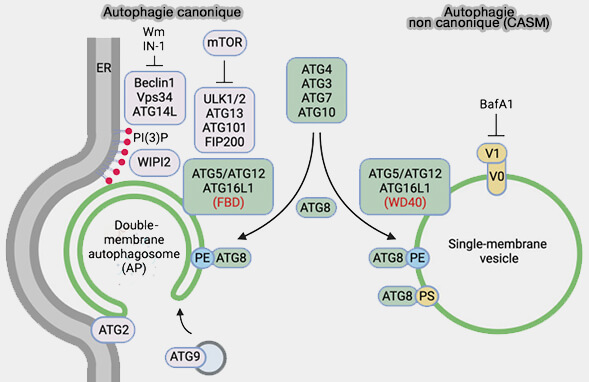
(Figure : vetopsy.fr d'après Durgan et Florey)
b. Récemment, une attache alternative d'ATG16L1, i.e. TECPR1, a été impliquée dans une autophagie dite aussi non canonique ( autophagie non canonique par TECPR1).
autophagie non canonique par TECPR1).
3. Ces deux autophagies impliquent des processus complètement différents, même si elles parviennent toutes deux à la lipidation de LC3.
La CASM modifie l’équilibre ionique et le pH de la vésicule endolysosomale cible grâce à une interaction avec la V-ATPase ( mécanisme général de la CASM).
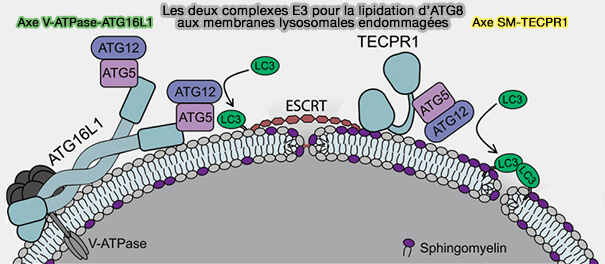
(Figure : vetopsy.fr d'après Corkery et Wu)
Vue d'ensemble de la CASM
1. La CASM implique les protéines ATG dans une gamme plus large de processus de dégradation que l'autophagie canonique par des liens partagés avec le lysosome.
- Sans avoir besoin d'initier la formation de novo d'autophagosome, l'autophagie non canonique est indépendante d'une grande partie des engins d'autophagie en amont, comme le complexe ULK/Atg1, i.e. ULK1/2, FIP200, ATG13, et ATG9, WIPI2 et ATG14.
- Cependant, les deux systèmes de conjugaison de type ubiquitine de base de la macroautophagie canonique, i.e le système ATG12, i.e. ATG4, ATG7, ATG3 et ATG12 et ATG16L1 et le système ATG8, i.e. ATG12, ATG7, ATG10 et ATG5, qui soutiennent la lipidation ATG8 dans les membranes, sont essentiels pour la CASM.
Remarque : les ATG sont aussi impliquées dans des processus non autophagiques (Autophagy-Independent Functions of the Autophagy Machinery 2019).

Nous ne pouvons pas développer toutes les voies de la CASM, mais en esquisser les points essentiels. Vous pouvez lire : Many roads lead to CASM: Diverse stimuli of noncanonical autophagy share a unifying molecular mechanism (2022), qui a servi à l'écriture de tous les chapitres sur la CASM, et vous reporter aux articles de la bibliographie.
3. La CASM, de plus en plus étudiée, sollicite des activateurs et des processus en constante expansion :
- les processus d'hétérophagie, i.e. phagocytose, endocytose, macropinocytose et entose,
- l'infection virale et d'autres facteurs pathogènes,
- la signalisation TRPML1 et STING,
- la neurodégénérescence,
- le cancer ( LDELS)
- la vision, par phagocytose et dégradation des segments externes des photorécepteurs par l’épithélium pigmentaire rétinien (Non-canonical Autophagy Promotes the Visual Cycle 2013),
- les traitements médicamenteux,
- la conservation évolutive (Evolutionary conservation and metabolic significance of autophagy in algae 2024).
Mécanisme général
PE versus PS
La CASM implique la lipidation d'ATG8 à la phosphatidyléthanolamine (PE), i.e. ATG8-PE, mais aussi à la phosphatidylsérine (PS), ATG8-PS, comme, par exemple, dans la phagocytose associée à LC3 (LAP).
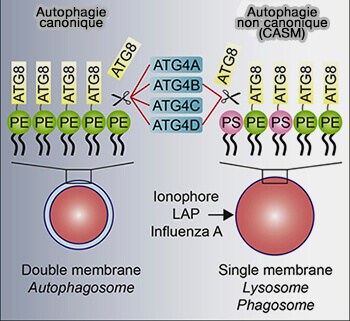
(Figure : vetopsy.fr d'après Tan et Finkel)
1. Dans le cas de la lipidation ATG8-PS, le mécanisme sous-jacent à la spécificité cellulaire n'est pas entièrement compris, mais la composition physiologique du pH et des phospholipides peut interdire cette lipidation alternative (Non-canonical autophagy drives alternative ATG8 conjugation to phosphatidylserine 2021).
2. Les différentes isoformes d'ATG4, i.e. A, B, C et D, peuvent délipider les ATG8-PE (Delipidation of mammalian Atg8-family proteins by each of the four ATG4 proteases 2018)
- ATG4D délipide préférentiellement LC3B-PS et GABARAP-PS.
- ATG4B prend également en charge la déconjugation partielle de l'ATG8-PS, ce qui suggère que des conditions modifiées, telles que la courbure de la membrane et/ou la charge, peuvent permettre cette activité.
Mécanisme général
La CASM est dépendante :
- d'ATG16L1,
- de la V-ATPase, la pompe à protons, constituée par le secteur transmembranaire V0 et le secteur cytosolique. V1.
La V-ATPase est étudiée dans un chapitre spécial.
1. L'augmentation de l'engagement de V0–V1 de la V-ATPase est suffisante pour pour l'activation de la CASM.
a. Après liaison de VO-V1, la V-ATPase récrute ATG16L1 via K490 pour provoquer la lipidation de LC3 ((The WD40 domain of ATG16L1 is required for its non‐canonical role in lipidation of LC3 at single membranes 2018).
b. Par exemple, au cours de la phagocytose associée à LC3 (LAP), la production de ROS dépendante de NOX2, une NADPH oxydase, consomme des protons phagosomaux d’une manière sensible au DPI et au GSK.
La modification du pH des phagosomes entraîne l’interaction entre la V-ATPase et son interaction avec ATG16L1, qui dirige ensuite la lipidation d’ATG8 vers les phagosomes (V-ATPase is a universal regulator of LC3-associated phagocytosis and non-canonical autophagy 2022).
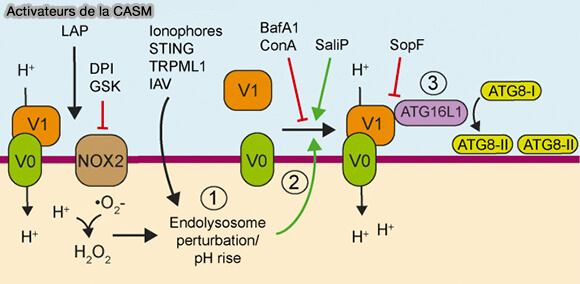
(Figure : vetopsy.fr d'après Xu et coll)
2. La bafilomycine A1 (BafA1) est un inhibiteur de la V-ATPase, empêchant l'association entre V0 et V1, qui augmente le pH lysosomal, perturbe la dégradation lysosomale et bloque le flux autophagique.
- En présence de BafA1, l’autophagie canonique se poursuit et les autophagosomes s’accumulent, produisant des niveaux accrus d’ATG8 lipidés provenant de cette voie.
- En revanche, BafA1 inhibe puissamment la lipidation ATG8 associée aux CASM, en réponse à un large éventail de stimuli.
La sensibilité BafA1 est une caractéristique déterminante de l’autophagie non canonique, qui peut différencier cette voie de la macroautophagie.
3. Certains agents pathogènes bloquent spécifiquement les CASM pour prospérer dans les compartiments de l’hôte.
La protéine SopF de Salmonella Typhimurium bloque l’interaction entre l'ATG16L1 et la V-ATPase pour abolir les MAEC (A Bacterial Effector Reveals the V-ATPase-ATG16L1 Axis that Initiates Xenophagy 2019).
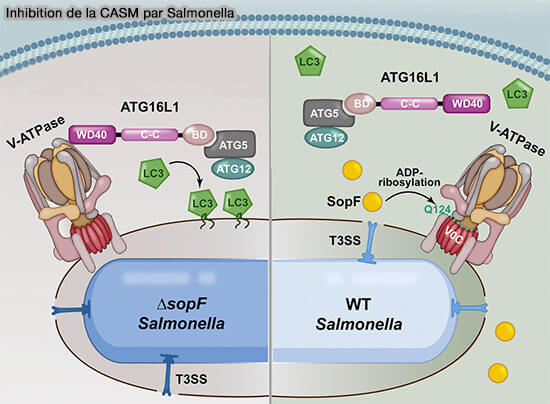
(Figure : vetopsy.fr d'après Xu et coll)
Dans l’autophagie canonique et non canonique, le ciblage membranaire du complexe E3 est médié par ATG16L1.
Différentes fonctions de la CASM
Biologie cellulaire et moléculaireConstituants de la celluleSystème endomembranaireRéticulum endoplasmiqueAppareil de GolgiEndosomesLysosomesPeroxysomesAutophagieMacroautophagieMicroautophagieAutophagie chaperonnes (CMA)Autophagie non canoniqueProtéines ATGSystème UPS (Ubiquitine-Protéasome)Transport membranaireTrafic vésiculaireFusion/fission membranaireEndocytoseVoie sécrétoireMoteurs moléculairesVoies de signalisation