


Neurophysiologie : synapse
Vésicules synaptiques : cycle vésiculaire
Organisation par condensats présynaptiques
- Neurophysiologie
- Cellules gliales et névroglie
- Neurones
- Potentiels membranaires
- Circuits neuronaux
- Neurotransmetteurs
- Récepteurs membranaires
- Synapses
{kind=link}

Les vésicules synaptiques et plusieurs protéines présynaptiques forment des condensats biomoléculaires dynamiques qui contribuent à l’organisation du bouton synaptique et à la régulation du cycle vésiculaire.
Les vésicules synaptiques et plusieurs protéines associées à la zone présynaptique participent à des condensats biomoléculaires dynamiques qui contribuent à l’organisation du compartiment présynaptique.
Ces condensats reposent sur des interactions multivalentes impliquant notamment des protéines riches en régions intrinsèquement désordonnées, comme la synapsine, qui favorisent le regroupement des vésicules et leur mobilité au sein des différents pools vésiculaires.

Les principes physico-chimiques de la séparation de phase liquide-liquide (LLPS) sont étudiés dans un chapitre spécifique.

(Figure : vetopsy.fr d'après Sansevrino et coll)
Condensats et organisation des vésicules synaptiques
Les terminaux nerveux contiennent plusieurs centaines de vésicules synaptiques (VS) qui sont étroitement regroupées dans le bouton présynaptique.
Des travaux récents suggèrent que ce regroupement repose en partie sur des condensats biomoléculaires dynamiques, dans lesquels les vésicules constituent une phase dispersée au sein d'une matrice protéique riche en protéines intrinsèquement désordonnées.
1. Les vésicules synaptiques présentent des propriétés compatibles avec une organisation de type phase fluide, dans laquelle elles peuvent se déplacer tout en restant confinées dans le bouton présynaptique.
Le regroupement des VS est notamment indépendant des protéines de la zone active (ZA) , puisque la suppression de certains composants de cette zone n'abolit pas l'accumulation des vésicules en amont (How to make an active zone: unexpected universal functional redundancy between RIMs and RIM-BPs 2016).
2. Malgré leur regroupement dans différents pools, les vésicules synaptiques restent hautement mobiles, ce qui permet leur recrutement rapide vers la membrane plasmique lors de l’activation neuronale (Subdiffractional tracking of internalized molecules reveals heterogeneous motion states of synaptic vesicles 2016).
Cette dynamique est compatible avec un modèle de condensat vésiculaire plutôt qu’avec un simple réseau d’échafaudage protéique.
Protéines impliquées dans l’organisation des condensats synaptiques
Plusieurs protéines des vésicules synaptiques (VS) sont à l'origine des condensats pour intervenir dans différentes fonctions.
Synapsine et regroupement des VS
1. La synapsine, par ses domaines D et E intrinsèquement désordonnés (IDR), suffit à induire la formation de micro-domaines de VS au sein du bouton synaptique, pour regrouper les vésicules lipidiques comme dans le pool de réserve ( synapsine et regroupement des VS).
synapsine et regroupement des VS).
Cela n'exclut pas un rôle supplémentaire par les interactions protéine-protéine comme avec :
- l'intersectine,
- l'α-synucléine.

(Figure : vetopsy.fr d'après Zhang et coll)
Condensat et endocytose synaptique
1. Dans l'endocytose, plusieurs protéines de courbure reposent sur des interactions faibles et de type liquide pour favoriser l'assemblage de gouttelettes de protéines ( LLPS et endocytose).
Ce serait le cas de :
- Eps15 et Fcho1/2,
- l'endophiline, comme aussi dans le cas de l'endocytose rapide endophiline-dépendante (FEME),
- la dynamine 1 par son interaction avec la syndapine 1 pour former des condensats moléculaires pour la médiation de l'endocytose ultrarapide (UFE) au niveau des synapses ( dynamine et UFE).
2. Les interactions cohésives au sein des condensats et les tensions interfaciales entre les condensats, les membranes et le cytosol pourraient contribuer à l'invagination membranaire au cours de l'endocytose dépendante et indépendante de l'actine.
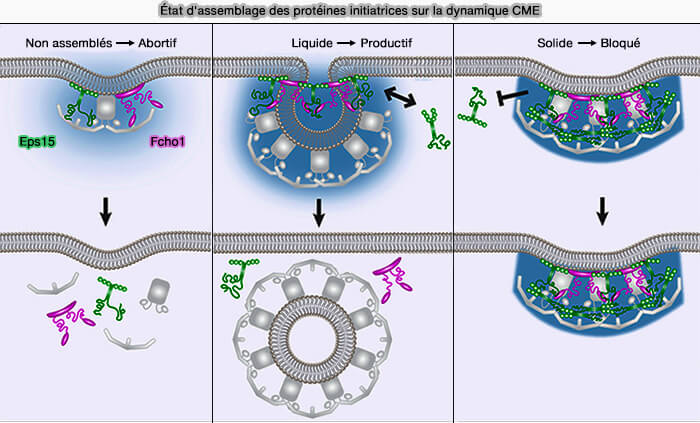
(Figure : vetopsy.fr d'après Day et coll)
Condensats de la zone active
Certaines protéines d’échafaudage présynaptique, notamment RIM et RIM-BP, peuvent former des condensats par séparation de phase liquide-liquide.
Ces assemblages contribuent à l’organisation de la zone active en regroupant les canaux calciques voltage-dépendants (CaV/VGCC) à proximité des sites de fusion vésiculaire.
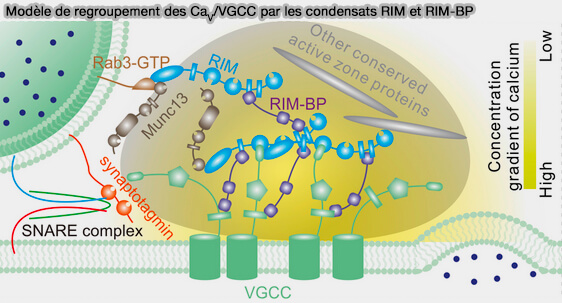
(Figure : vetopsy.fr d'après Wu et coll)
Retour au cycle vésiculaire
NeurophysiologeCellules gliales et névroglieNeuronesPotentiels membranairesCircuits neuronauxNeurotransmetteursRécepteurs membranairesSynapsesStructure synaptiqueNeurotransmissionVésicules synaptiques (VS)Pools synaptiquesRôles du calcium dans l'électrosécrétionCycle synaptiqueTranslocation et attache (tethering)Amarrage (docking) et amorçage (priming)Fusion et exocytoseRecyclage des VSDifférents types d'endocytoseAcidification et remplissage des VSRegroupement des VSPlasticité et régulation synaptique