1. Ainsi, les machines de conjugaison, et par extension les protéines GABARAP, sont importantes pour une fermeture efficace et une fission de la membrane autophagosomale interne et externe au bord du phagophore.
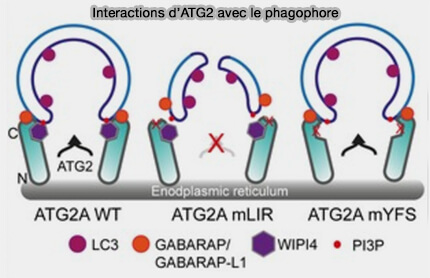
Interactions d'ATG2 avec le phagophore
(Figure : vetopsy.fr d'après Bozic et coll)
La manière dont elles contribuent à la fermeture est obscure, mais semble être spécifique à la sous-famille GABARAP.
Remarque : la déplétion en WIPI4 provoque également des structures ouvertes de phagophore, mais elles sont morphologiquement différentes de celles générées après la déplétion d'ATG2A/B2 ( rôle des WIPI).
Il n'est toujours pas clair de savoir si cette fonction est nécessaire à la fission de la membrane de phagophore, mais il a été proposé que les protéines GABARAP puissent combler une partie du pore de fission pour faciliter la fermeture des organites.
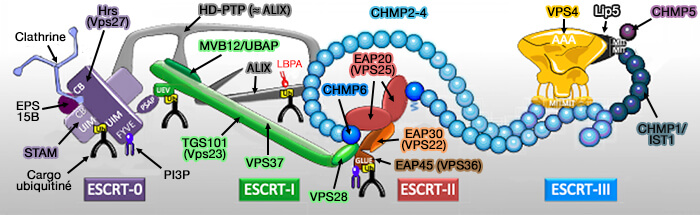
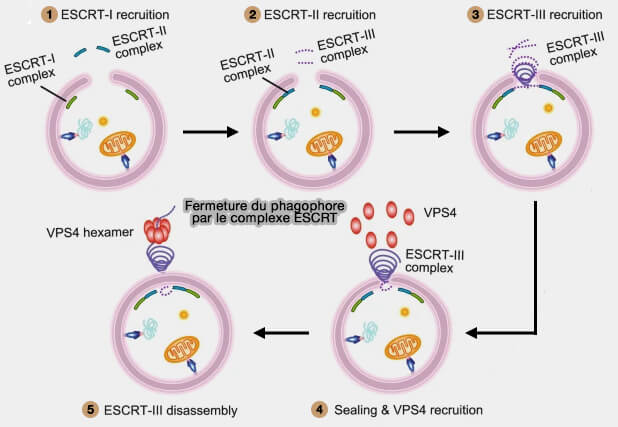
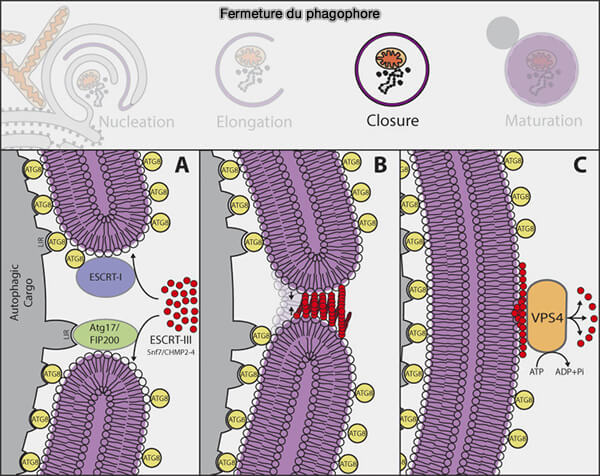
b. Vient ensuite, la dépolymérisation de l’ESCRT-III par Vps4 (Vacuolar Protein Sorting 4) qui contient un domaine catalytique AAA+ ATPase essentiel au fonctionnement des complexes ESCRT.
Remarque : l’inhibition de la fermeture de la membrane par la déplétion de CHMP2A ou l’inhibition de VPS4 accumule VPS37A sur le phagophore.
Ces observations suggèrent que VPS37A coordonne le recrutement d’un ensemble unique de composants de la machinerie ESCRT pour la fermeture du phagophore dans les cellules de mammifères.
Les ESCRT ne peuvent favoriser que la toute dernière étape de scission d’un col (ou d’une ouverture) membranaire très rétréci, ce qui implique que d’autres mécanismes encore inconnus doivent exister pour resserrer le bord du phagophore à ce stade.
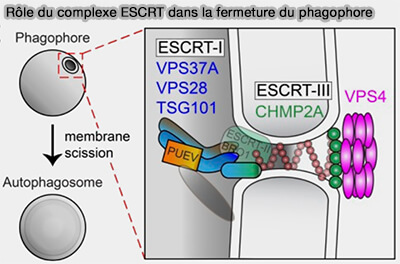
Fermeture du phagophore par le complexe ESCRT
(Figure : vetopsy.fr d'après Li
et coll)
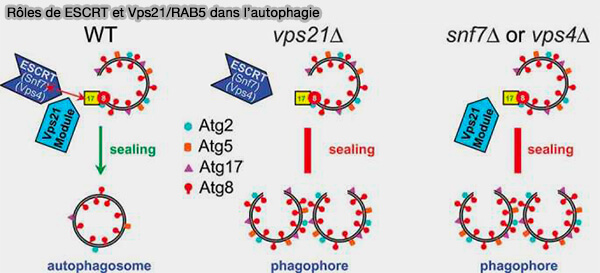
Rôles de ESCRT et Vps21/RAB5 dans l’autophagie
(Figure : vetopsy.fr d'après Zhou
et coll)
3. Le rôle essentiel des ESCRT dans la fermeture des autophagosomes est mis en évidence par le fait que la déplétion des ESCRT provoque l’accumulation d’autophagosomes ouverts et une altération de l’autophagie.
Toutefois, il semble que le flux autophagique ne soit pas complètement aboli, ce qui suggère qu’une partie des autophagosomes peut se fermer en l’absence d’une machinerie ESCRT intacte (Rab5-dependent autophagosome closure by ESCRT 2019).
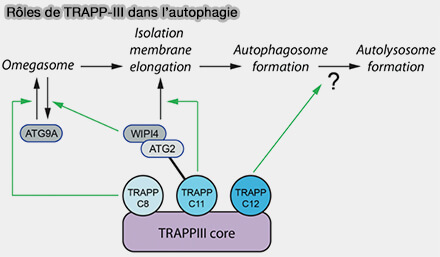
Complexe TRAPP-III et autophagie
(Figure : vetopsy.fr d'après Stanga et coll)
Le retard de dégradation de la membrane interne autophagosomale dans les cellules déficientes en conjugaison ESCRT ou ATG pourrait être très probablement la conséquence de l'acquisition de LAMP1 et/ou de lipides lysosomaux et pourrait expliquer l'accumulation d'autophagosomes observée dans les cellules déplétées en ESCRT (ESCRT-III Dysfunction Causes Autophagosome Accumulation and Neurodegeneration 2007).
En conséquence, les défauts dans les systèmes conjugués ou dans le transport lipidique à médiation Atg2 jusqu'au phagophore en croissance se traduisent par l'accumulation de membranes d'isolement anormales, de vésicules préautophagosomales positives LC3 et d'autophagosomes incomplets.
2. La syntaxine17 (STX17), impliquée dans la fusion des autophagosomes avec les lysosomes, n’est recruté semble-t-il que quand l’autophagosome a été scellé.
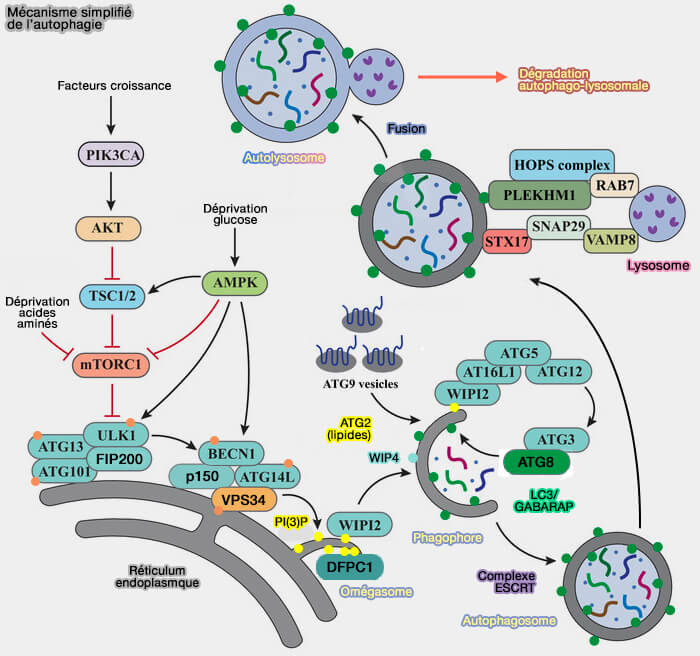
Mécanisme simplifié de l'autophagie
(Figure modifiée : vetopsy.fr d'après Lu et coll)





 rôle d'ATG2 dans la fermeture de l'autophagosome et partenaires d'Atg2).
rôle d'ATG2 dans la fermeture de l'autophagosome et partenaires d'Atg2).