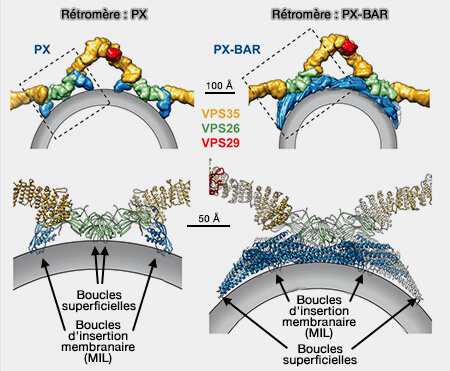
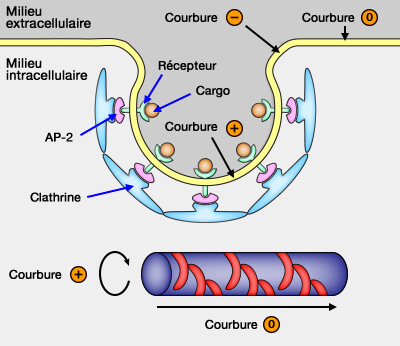
Le signe de la courbure membranaire est tout à fait arbitraire : il se réfère à un observateur qui serait placé dans le cytoplasme.
Les membranes planes n'ont pas de courbure (courbure 0).
Les invaginations, qui pénètrent dans le cytosol, possèdent une courbure positive.
Les saillies hors du cytosol ont une courbure négative.
Remarque : pour un tubule, formé et contraint par des protéines, la courbure est positive le long de la face latérale du cylindre et nulle le long de l'axe tubulaire.
Les domaines BAR forment généralement des homodimères en forme de banane ou de croissant, où chaque protomère est constitué d'une structure enroulée antiparallèle à trois hélices.
En raison de sa forme incurvée, le domaine BAR est capable de détecter la courbure de la membrane, i.e. de se lier préférentiellement aux membranes incurvées, i.e.
Ainsi, la forme concave du dimère à domaine BAR est chargée positivement et interagit avec la monocouche cytoplasmique chargée négativement des biomembranes.
La plupart des protéines à domaine BAR contiennent également des domaines auxiliaires impliqués dans les interactions avec d'autres protéines ou avec des membranes ( plus bas).
Les facteurs de cette homodimérisation sont largement inconnus, mais permettent la régulation de ces protéines.
En effet, cette dimérisation est essentielle pour courber la surface membranaire.
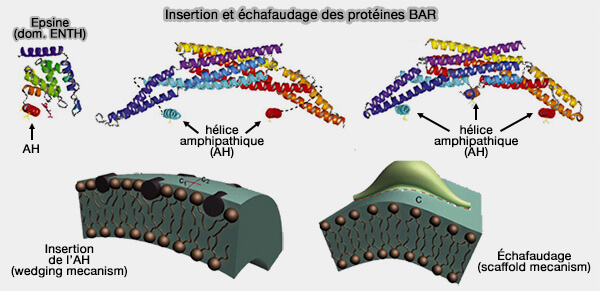
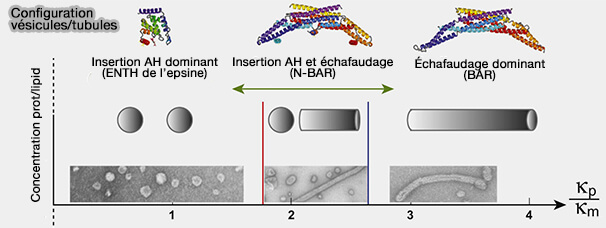
Insertion de l'hélice amphipathique et échafaudage des protéines BAR
(Figure : vetopsy.fr d'après Boucrot et coll)
2. Les domaine BAR peuvent employer :
des mécanismes d'échafaudage (scaffolding mechanism) : des résidus du dimère chargés positivement interagissent avec la bicouche de lipides chargés négativement de la membrane (Structural Basis of Membrane Invagination by F-BAR Domains 2008) et contraignent la forme de la membrane en la stabilisant,
des mécanismes d'enfoncement d'un " coin " (wedging mechanism), comme l'hélice amphipathique des N-BAR, qui déplace les lipides de la couche interne pour " plier " la membrane vers le domaine BAR, et entraine le plus souvent une fission membranaire ( fission passive).
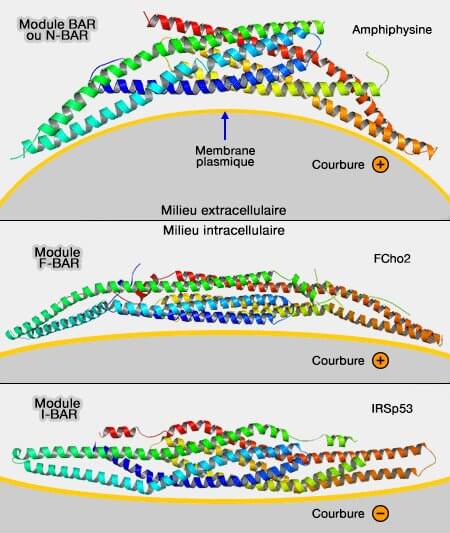
Les domaines BAR classiques ont une affinité pour les membranes fortement courbées positivement et sont contenus dans plusieurs protéines.
Les N-BAR sont le plus souvent inclus dans les modules BAR classiques, en opposition avec les F-BAR et les I-BAR.
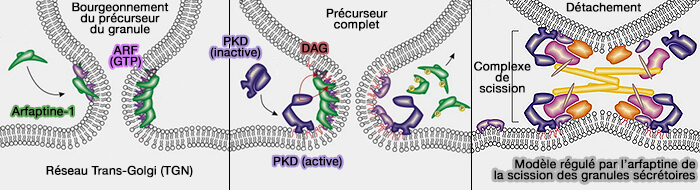
1. Les arfaptines se lient à la petite GTPase Arf1 pour former un dimère allongé en forme de croissant avec une superhélice (coiled-coil) de trois hélices α impliquée dans le bourgeonnement :
l'hélice amphiphile N-terminale (H0), qui assure la liaison à la membrane,
l'hélice amphiphile insérée dans l'hélice 1 (H1), qui s'insère dans la membrane,
le domaine structurel, en forme de croissant formé par sa dimérisation, qui coopère avec H1 pour courber la membrane.
2. Les domaines N-BAR possèdent une hélice amphipathique N-terminale qui précède le domaine BAR qui s'enfonce peu profondément dans les membranes.
Cette hélice, comme l'hélice α0 du domaine ENTH de l'epsine, s'insère entre les couches de la membrane plasmique pour l'incurver.
Différents modules (dimères) BAR
(Figure : vetopsy.fr d'après endocytosis.org)
Si on retire les résidus 1 à 26 de la partie N-terminale, cette hélice ne se forme plus. Le module BAR stabilise l'incurvation.
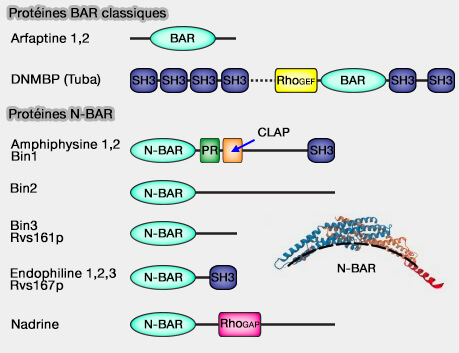
un domaine N-BAR qui s'homodimérise pour former un module BAR avec son hélice amphipathique,
un domaine central CLAP (CLathrin and AdaPtor binding domain),
un domaine SH3 C-terminal qui se lie avec le domaine PRD (PR ou PRD : riche en proline) de la dynamine (ProSerArgProAsnArg).
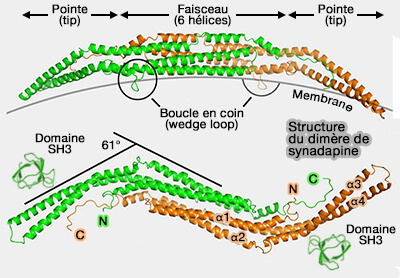
a. Chaque monomère est un enroulement en spirale de trois longues hélices α, formant un faisceau de six hélices autour de l'interface du dimère, en forme de banane.
La courbure du dimère est due d'une part au croisement des monomères et d'autre part aux anomalies dans les hélices 2 et 3.
L'interface entre les monomères est en grande partie hydrophobe.
Bin1 (Bridging integrator 1), acteur clé dans le contrôle de la courbure, de la mise en forme et du remodelage de la membrane plasmique et requis dans les cellules musculaires pour la formation des tubules en T et lors de l'endocytose rapide endophiline-dépendante (FEME).
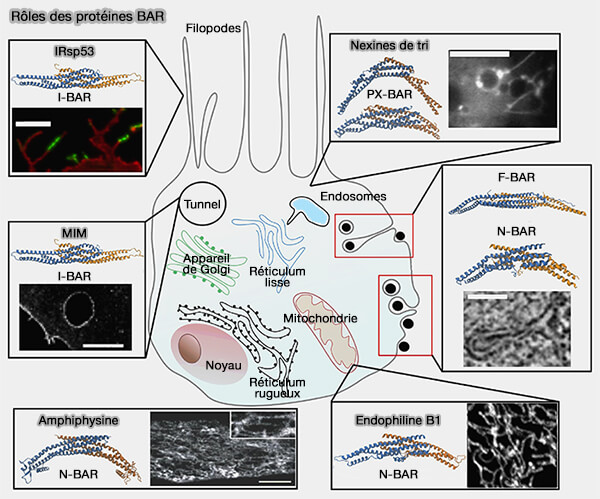
Quelques protéines à domaine BAR classique et N-BAR
(Figure : vetopsy.fr) Bin2 (Bridging integrator 2) favorise la motilité et la migration cellulaires, probablement via son interaction avec la membrane cellulaire et avec les protéines du podosome qui interviennent dans l'interaction avec le cytosquelette.
la nadrine (Rho GTPase activating protein 17), GTPase neuronale impliquée dans l'exocytose et dans le maintien de la jonction serrée en régulant l'activité de CDC42, jouant ainsi un rôle central dans la polarité apicale des cellules épithéliales.
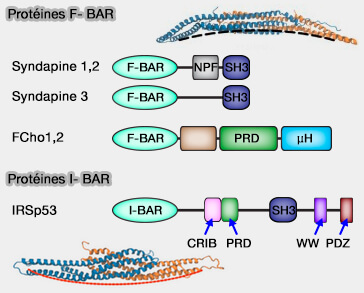
Modules F-BAR
Les modules F-Bar (FCH-BAR, Fes/CIP4 Homology-BAR ou EFC/F-BAR pour FCH Extended Homology) reconnaissent différentes courbures positives, mais moins profondes que les modules précédents (F-BAR domain proteins 2010).
les N-BAR nadrine et oligophrénine et les protéines du
les F-BAR srGAP1-4, et HMHA1.
b. Un domaine RhoGEF est présent dans DNMBP (Tuba).
c. Des domaines ArfGAP, i.e. activateurs des petites GTPases Arf, sont présents dans la centaurine et ASAP1.
Tous ces domaines indiquent un lien très étroit entre les protéines BAR, les petites GTPases connues en tant que régulateurs du cytosquelette d'actine.
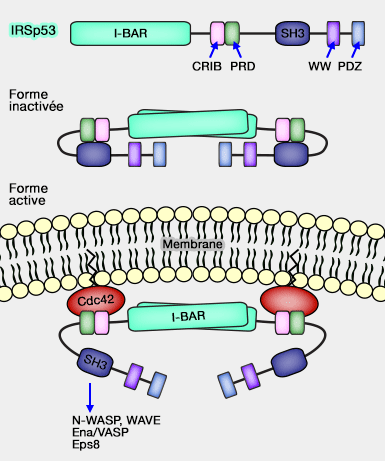
IRSp53 et son activation
(Figure : vetopsy.fr)
Son domaine C-terminal, riche en proline contribue à son auto-inhibition par sa liaison avec son domaine SH3.
Ce domaine se lie à Cdc42 pour le déplier et exposer le domaine SH3.






 plus bas).
plus bas).