


Autophagie
Macroautophagie
Autophagie sélective de la levure
Atg11
- Vue d'ensemble du système endomembranaire
- Autophagie
- Système UPS (Système UPS (Ubiquitine-protéasome))
- Fusion et fission membranaire
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

Chez la levure, Atg11 a une architecture en domaines coiled coil (superhélices) qui permet sa dimérisation parallèle, ce qui expliquerait la variation de l'organisation du PAS pour l'autophagie sélective.
Vue d'ensemble de l'autophagie sélective de la levure
Les protéines d’échafaudage et des composants d’attache membranaire, comme Atg11 et Atg17, sont nécessaires pour la formation d’autophagosomes chez la levure.
Ces protéines servent de plaques tournantes ou de plates-formes pour le recrutement de la machinerie autophagique de base.
1. Atg17, sous-unité du complexe kinase Atg1, recrute et attache les vésicules donneuses d’Atg9 dans la macrophagie non sélective ( interactions Atg17/Atg9).
interactions Atg17/Atg9).
Le complexe kinase Atg1 correspondant induit la formation d’autophagosomes non sélectifs (Organization of the Pre-autophagosomal Structure Responsible for Autophagosome Formation 2008).
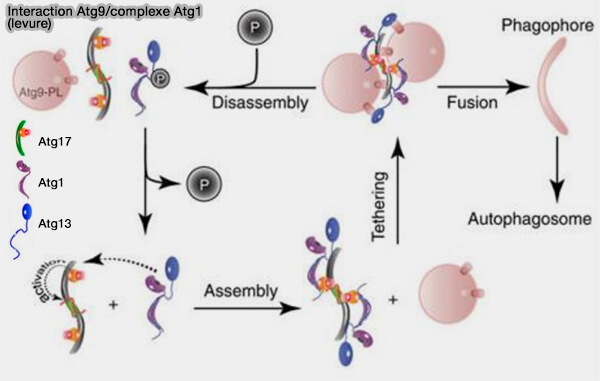
(Figure : vetopsy.fr d'après Rao et coll)
2. Alors qu'Atg17 est suffisant pour conduire à l’autophagie non sélective, l’implication d’Atg11 est nécessaire pour l’autophagie sélective, qui, en plus de jouer une fonction similaire à Atg17, joue un rôle essentiel dans l’organisation et l’activation du PAS sélectif spécifique à l’autophagie (PAS sélectif) en interagissant avec les complexes récepteur sélectif (SAR)/cargo.
Atg11
Atg11 est la deuxième plus grande Atg, i.e.1178 résidus, après Atg2.

Vous pouvez lire : Mechanistic insights into the role of Atg11 in selective autophagy (2019).
Structure d'Atg11
1. Malgré sa taille, la seule caractéristique structurelle est constituée par ses quatre domaines coiled-coil (CC1 à 4) qui sont répartis dans sa séquence.
- Les CC constituent l'un des principaux motifs d'oligomérisation dans les protéines (Coiled‐coils: The long and short of it 2016).
- Les CC naturellement présentes ont entre deux et cinq hélices α amphipathiques qui se tordent les unes autour des autres pour former une superhélice.
- Les CC peuvent être disposés selon des orientations parallèles ou antiparallèles.
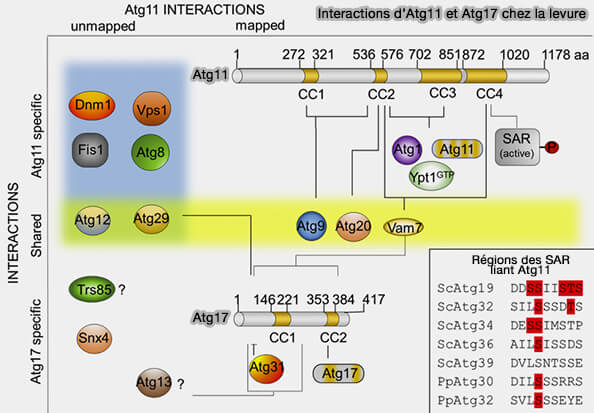
(Figure : vetopsy.fr d'après Zientara-Rytter et Subramani)
2. CC2 et le CC3 sont des régions nécessaires à la dimérisation d'Atg11 au PAS, mais CC4 est également essentiel pour ce processus et pour les interactions avec Atg1 et Ypt1/Rab1 (Receptor-bound targets of selective autophagy use a scaffold protein to activate the Atg1 kinase 2015 et Two Independent Pathways within Selective Autophagy Converge to Activate Atg1 Kinase at the Vacuole 2016).
a. Dans le PAS sélectif, Atg11 semble être une composante à part entière du complexe de kinases Atg1, avec lequel il interagit par le biais de ses régions CC2.
- Atg11 interagit avec le complexe Atg17/Atg31/Atg29, qui relie les étapes de la sélection des cargos à l'initiation de la biogenèse de l'autophagosome.
- L'Atg11 interagit également avec Atg20, qui est perturbé lorsque CC2 de l'Atg11 est supprimé, et avec Atg29, qui doit être phosphorylé pour interagir avec Atg11 (Atg29 phosphorylation regulates coordination of the Atg17-Atg31-Atg29 complex with the Atg11 scaffold during autophagy initiation 2013).
- L'Atg13, une autre composante du complexe Atg1, régule l'interaction entre le complexe Atg1 et l'Atg11.
b. CC2, et peut-être CC1, interagit avec Atg9.
L'initiation du phagophore dépend du recrutement et de la fusion des vésicules Atg9 ( trafic d'Atg9).
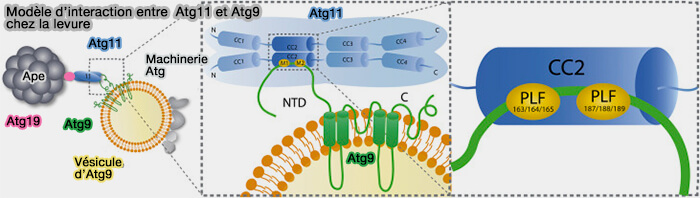
(Figure : vetopsy.fr d'après Coudeville et coll)
3. CC4 est responsable de la liaison aux récepteurs de l'autophagie sélective ou SAR (Atg11 Links Cargo to the Vesicle-forming Machinery in the Cytoplasm to Vacuole Targeting Pathway 2005).
Le CC4 est également conservé dans le domaine C-terminal de la protéine de mammifère FIP200/RB1CC1, dont la fonction est similaire à celle d'Atg17, suggérant la conservation évolutive du rôle du domaine CC4 dans l'autophagie.
Atg11 versus Atg17
1. Atg11 a une architecture de CC parallèles (Biophysical characterization of Atg11, a scaffold protein essential for selective autophagy in yeast 2017).
Contrairement à Atg11, Atg17 a des CC antiparallèles et forme des dimères en forme de S (Architecture of the Atg17 Complex as a Scaffold for Autophagosome Biogenesis 2012).

Ces différences architecturales entre Atg11 et Atg17 refléteraient la variation de l'organisation du PAS pour l'autophagie sélective et non sélective.
(Figure : vetopsy.fr d'après Zientara-Rytter et Subramani)
2. Dans le PAS d'autophagie sélective, la fixation de multiples dimères d'Atg11 à la surface du cargo par l'intermédiaire des récepteurs de l'autophagie sélective (SAR) pourrait suffire à adopter la forme caractéristique du phagophore, qui entoure et enferme la cargaison à l'intérieur de l'autophagosome.
3. Dans le PAS d'autophagie non sélective, où le cargo n'a pas de forme/structure conservée, l'assemblage du phagophore en forme de coupe nécessite des interactions tridimensionnelles répétées avec des complexes Atg17 en forme de S de 34 nm de long pour surmonter une barrière cinétique importante dans cette transition de forme de membrane (Scaffolding the cup-shaped double membrane in autophagy 2017).
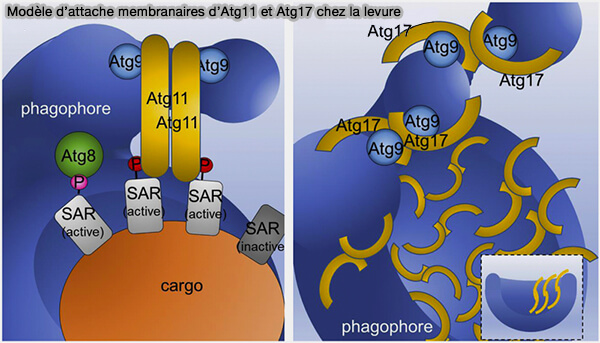
Modèle d'action d'Atg11
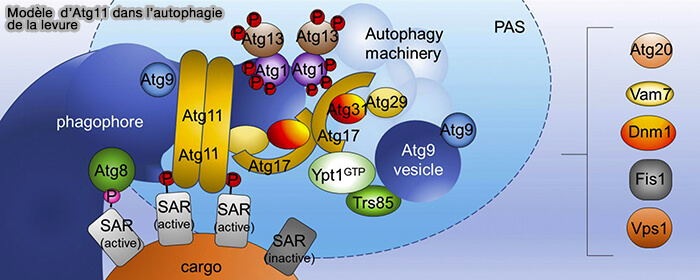
(Figure : vetopsy.fr d'après Zientara-Rytter et Subramani)
1. Atg11 initie l’autophagie sélective et coordonne le passage à l’autophagie non sélective en recrutant Atg9 (Atg11 tethers Atg9 vesicles to initiate selective autophagy 2019).
- Atg11 est activé par le cargo par une interaction directe avec les récepteurs de l'autophagie sélective (SAR).
- Atg11 activé dimérise par son domaine C-terminal et, par exemple, par son interaction avec le récepteur mitochondrial Atg32, et attache les vésicules Atg9, ce qui conduit à la nucléation des phagophores à proximité directe du cargo.
- En outre, ATG11 se lie plus efficacement à Atg9 (60%) qu'à Atg17 (10%).
2. La dénutrition régule l’activité des deux facteurs d’attache en initiant la dégradation d’Atg11 et en diminuant sa concentration d'un facteur 10 à 100, alors que la concentration d'Atg17 activé reste constante.
Cela permet à Atg17 de séquestrer et d’attacher les vésicules Atg9 indépendamment du cargo pour nucléer des phagophores non sélectifs.
3. Ensuite, la fusion homotypique de ces vésicules donne naissance au phagophore (Atg9 vesicles are an important membrane source during early steps of autophagosome formation 2012).
Modèle actuel d'autophagie sélective chez les mammifères
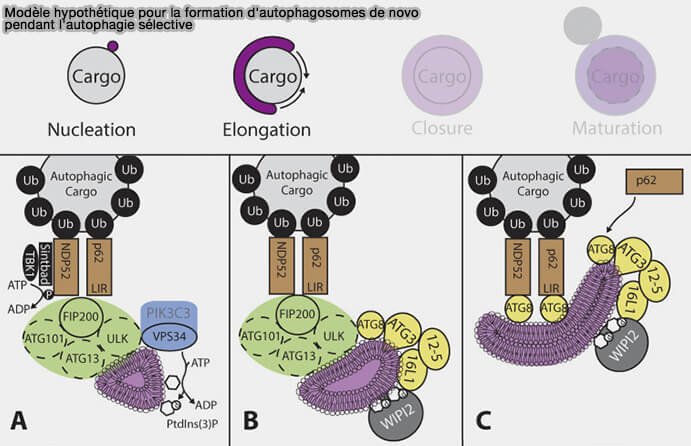
(Figure : vetopsy.fr d'après Melia et coll)
Biologie cellulaire et moléculaireConstituants de la celluleSystème endomembranaireRéticulum endoplasmiqueAppareil de GolgiEndosomesLysosomesPeroxysomesAutophagieMacroautophagieMicroautophagieAutophagie chaperonnes (CMA)Autophagie non canoniqueProtéines ATGSystème UPS (Ubiquitine-Protéasome)Transport membranaireTrafic vésiculaireFusion/fission membranaireEndocytoseVoie sécrétoireMoteurs moléculairesVoies de signalisation