


Autophagie
Protéines ATG (AuTophaGy-related)
ATG16L
- Vue d'ensemble du système endomembranaire
- Autophagie
- Système UPS (Système UPS (Ubiquitine-protéasome))
- Fusion et fission membranaire
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

L'ATG16L dimérique forme un complexe avec ATG12-ATG5 qui intervient comme une E3 ligase pour la lipidation d'ATG8, aussi bien dans l'autophagie canonique que dans la CASM (Conjugation of ATG8 to Single Membranes).
La lipidation d'ATG8 nécessite deux systèmes de type ubiquitine :
- le système ATG12,
- le système ATG8.
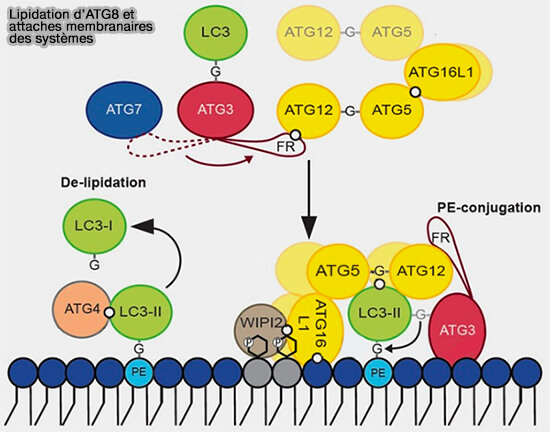
(Figure : vetopsy.fr d'après Lystag et Simonsen)
ATG16L comprend plusieurs isoformes :
- ATG16L1,
- ATG16L2.
ATG16L1
Structure d'ATG16L
La structure d'ATGL1 est décrite dans la figure ci-dessous.
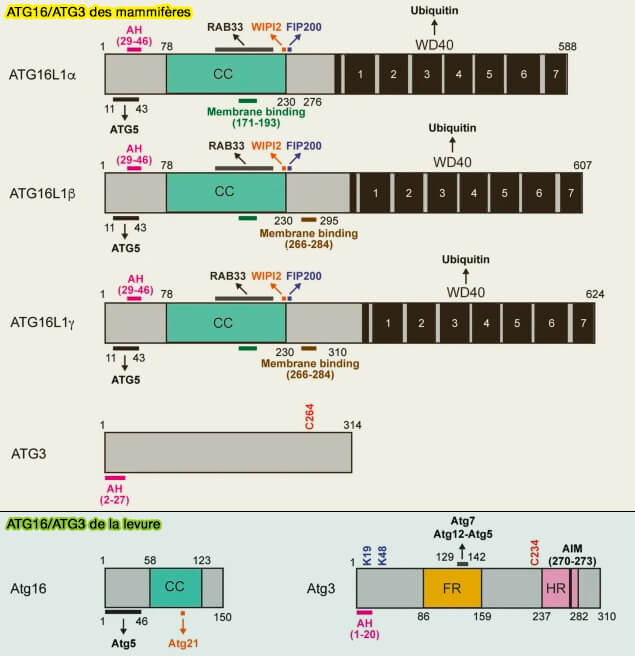
(Figure : vetopsy.fr d'après Nishimura et Tooze)
Fonctions d'ATG16L
Dans l'autophagie canonique
1. Dans la macroautophagie canonique, ATG16L1 n'est pas nécessaire à la réaction de lipidation d'ATG8, mais sa liaison membranaire détermine le site de la lipidation de l’ATG8 au niveau du phagophore dépendant de PI(3)P.
a. Lors de déprivation d’acides aminés ou de glucose, le recrutement membranaire de ATG16L1 implique sa liaison à WIPI2 ( liaison ATG16L1/WIPI2).
liaison ATG16L1/WIPI2).
Son interaction avec WIPI2 (8 sur la figure ci-dessous) assure la liaison à PI(3)P.
b. ATG16L1 interagit avec ATG5 et la membrane par deux hélices (Distinct functions of ATG16L1 isoforms in membrane binding and LC3B lipidation in autophagy-related processes 2019) :
- avec Atg5 par son hélice N-terminale, i.e hélice 1, 13–28 (4),
- avec la membrane par une hélice amphipathique (AH), i.e. hélice 2, 29-46, requise pour la lipidation des protéines de la famille LC3 (7).
Les résidus de cette hélice s’intègrent partiellement dans la membrane, afin d’amener le conjugué ATG5-ATG12 près de la membrane, permettant à ATG3-A2TG8 d’atteindre le co-substrat, i.e. PE/PS ((Membrane curvature sensing and stabilization by the autophagic LC3 lipidation machinery 2022).
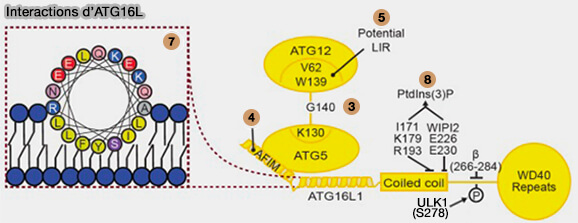
(Figure : vetopsy.fr d'après Lystag et Simonsenl)
Remarque : la lipidation d'ATG8 pourrait se produire sur des membranes non autophagiques, mais elle est efficacement corrigée par la déconjugaison médiée par ATG4 qui peut être régulée par Atg1 et ULK1 ( régulation de l'ATG8 par ATG4).
2. Cependant, ATG16L1 peut cibler d'autres membranes indépendamment de WIPI2.
a. La liaison membranaire à travers l’insert spécifique de l’isoforme β, i.e. 266-284, est importante pour la lipidation de la protéine ATG8 des endosomes/lysosomes endommagés.
- Cette région β est essentielle pour cette lipidation indépendante du VPS34 du complexe PI3KC3-CI (Distinct functions of ATG16L1 isoforms in membrane binding and LC3B lipidation in autophagy-related processes 2019).
- L’insert spécifique à l’isoforme β est une zone fortement modifiée post-traductionnellement dans ATG16L1 et comprend un site de phosphorylation confirmé pour ULK1 sur S278.
c. Le domaine CC a une capacité intrinsèque à se lier aux lipides, ce qui pourrait améliorer l'interaction membranaire (Intrinsic lipid binding activity of ATG16L1 supports efficient membrane anchoring and autophagy 2019).
d. Le domaine WD intervient dans la CASM ( cf. plus bas).

Le complexe Atg16/ATG16L1 est recruté dans les membranes cibles par l'intermédiaire de partenaires à interaction multiple, y compris les protéines et les lipides qui peuvent être modifiés en réponse à un contexte physiologique.
Dans l'autophagie non canonique (CASM)
1. La CASM (Conjugation of ATG8 to Single Membranes), autophagie non canonique, est dépendante :
- d'ATG16L1,
- de la V-ATPase, la pompe à protons, constituée par le secteur transmembranaire V0 et le secteur cytosolique. V1.
La CASM est étudiée dans des chapitres spéciaux.
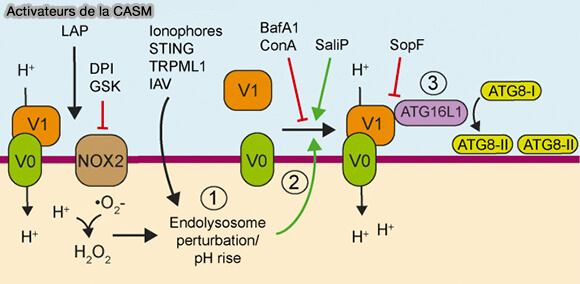
(Figure : vetopsy.fr d'après Xu et coll)
2. Le domaine WD d'ATG16L1 via K490 est recruté par la V-ATPase pour provoquer la lipidation de LC3 (The WD40 domain of ATG16L1 is required for its non‐canonical role in lipidation of LC3 at single membranes 2018).
ATG16L2
Peut-être dans le but d’augmenter le répertoire d’événements lipidateurs d’ATG8, les vertébrés ont développé deux partenaires complexes alternatifs pour ATG5-ATG12, i.e. :
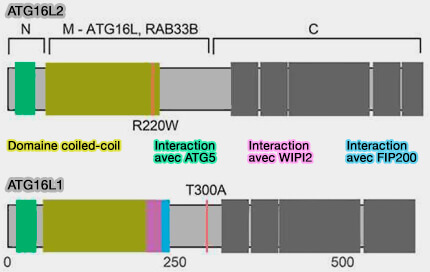
(Figure : vetopsy.fr d'après Don Wai Luu et coll)
1. ATG16L2 est similaire à ATG16L1 dans sa structure, mais diffère par son domaine coiled-coil (CC), en particulier dans la région 229-242 qui correspond à la liaison WIPI2 et FIP200 d'ATG16L1 (Distinct tissue-specific roles for the disease-associated autophagy genes ATG16L2 and ATG16L1 2019).
- Chez l’homme, ATG16L2 peut former un homodimère ou un hétérodimère avec ATG16L1.
- De plus, la région N-terminale ATG16L2 peut également se lier à ATG12-ATG5 avec une affinité similaire à ATG12-ATG5-ATG16L1.
2. ATG16L2 n’est pas recruté dans les phagophores et est principalement présent dans le cytosol (Atg16L2, a novel isoform of mammalian Atg16L that is not essential for canonical autophagy despite forming an Atg12–5-16L2 complex 2011).
- Atg16L2 est incapable de compenser la fonction d’Atg16L1 dans la formation d’autophagosomes.
- L’inactivation d’Atg16L2 endogène n’a pas affecté la formation d’autophagosomes, indiquant qu'Atg16L2 ne possède pas la capacité de médier l’autophagie canonique.
3. ATG16L2 pourrait entrer en compétition avec ATGL16L1 pour la formation de complexe avec ATG12-ATG5 et réduirait l'autophagie, ce qui serait important dans la xénophagie, mais les études doivent encore être affinées (ATG16L2 inhibits NLRP3 inflammasome activation through promoting ATG5-12-16L1 complex assembly and autophagy 2022).
- Par contre, ATG16L2 joue un rôle important dans la pathogenèse de divers cancers, maladies auto-immunes et troubles neurodégénératifs (The role of ATG16L2 in autophagy and disease 2022).
- Bon nombre des études actuellement disponibles ont identifié une association entre l’expression de ATG16L2 et la maladie, mais n’ont pas encore identifié directement si ou comment ATG16L2 est impliqué dans la maladie.
Retour à l'élongation du phagophore
Biologie cellulaire et moléculaireConstituants de la celluleSystème endomembranaireRéticulum endoplasmiqueAppareil de GolgiEndosomesLysosomesPeroxysomesAutophagieMacroautophagieMicroautophagieAutophagie chaperonnes (CMA)Autophagie non canoniqueProtéines ATGSystème UPS (Ubiquitine-Protéasome)Transport membranaireTrafic vésiculaireFusion/fission membranaireEndocytoseVoie sécrétoireMoteurs moléculairesVoies de signalisation