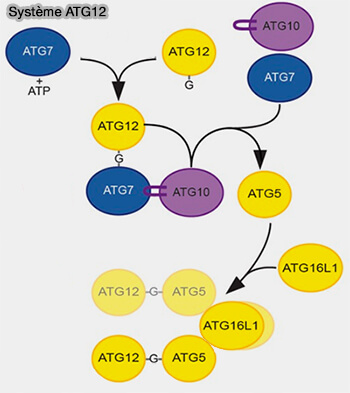
Lors de l'élongation du phagophore, la lipidation d'ATG8 implique deux systèmes de conjugaison covalente de type ubiquitine, le système ATG12 et le système ATG8.
La biogenèse des autophagosomes implique plusieurs processus :
Le phagophore doit s’allonger et s’incurver autour du fragment cytoplasmique à dégrader, pour finir par se refermer pour former l'autophagosome sous l'action de deux complexes de conjugaison de type ubiquitine représentés par Atg12 et Atg8/LC3.
L'élongation et la courbure membranaire impliquent :
deux systèmes de conjugaison de type ubiquitine, le système ATG12 et le système ATG8,
Le but ultime de ces réactions enzymatiques est la lipidation, i.e. liaison covalente avec la phosphatidyléthanolamine (PE), et donc l'ancrage des protéines de la famille Atg8 à la membrane de phagophore en croissance.
La phosphatidyléthanolamine (PE) est le deuxième phospholipide le plus abondant après la phosphatidylcholine (PC).
b. La lipidation d’ATG8 pourrait se produire sur des membranes non autophagiques, mais elle est efficacement corrigée par la déconjugaison médiée par ATG4 qui peut être régulée par Atg1 et ULK1 ( régulation de l'ATG8 par ATG4).
ATG16L1 peut cibler d'autres membranes indépendamment de WIPI2 comme dans la CASM (Conjugation of ATG8 to Single Membranes).
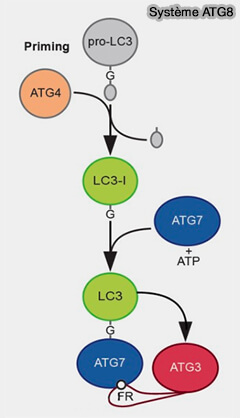
Système ATG8
Le système ATG8/LC3, dans le même temps, et sous l’influence du système ATG12, intervient dans l'élongation.
1. ATG8/LC3, une autre protéine de type ubiquitine, est d’abord synthétisée sous forme immature, i.e. pro-ATG8 ou pro-LC3, dont la région C-terminale est clivée par les enzymes de la famille ATG4 pour exposer un résidu de glycine et former ATG8-I ou LC3-I.
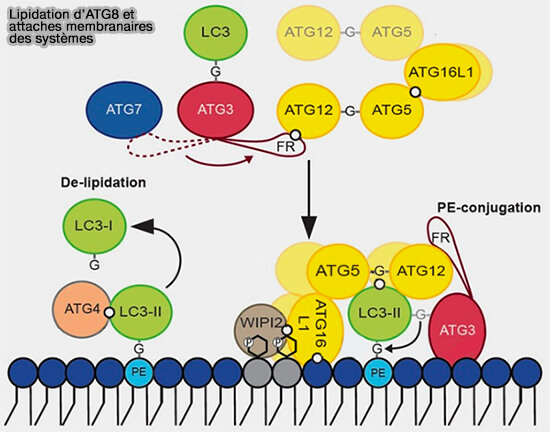
Le conjugué ATG12-ATG5 agit comme une enzyme de type E3 pour promouvoir la conjugaison ATG8-PE ( régulation allostérique d'Atg3).
3. Contrairement à la conjugaison irréversible de l’ATG12, l’ATG8-PE peut être à nouveau déconjuguée par l’ATG4 ( régulation de l'ATG8 par ATG4).
Lipidation d’ATG8 et attaches membranaires des systèmes
(Figure : vetopsy.fr d'après Lystag et Simonsen)
3. Les fonctions de l'ATG8, dans ce cas GABARAP, seraient :
une partie de l'approvisionnement en lipides des membranes,
le complexe final, i.e. ATG12-ATG5-ATG16L1-ATG3-ATG8, qui participe à la courbure membranaire ( fonctions d'ATG8).
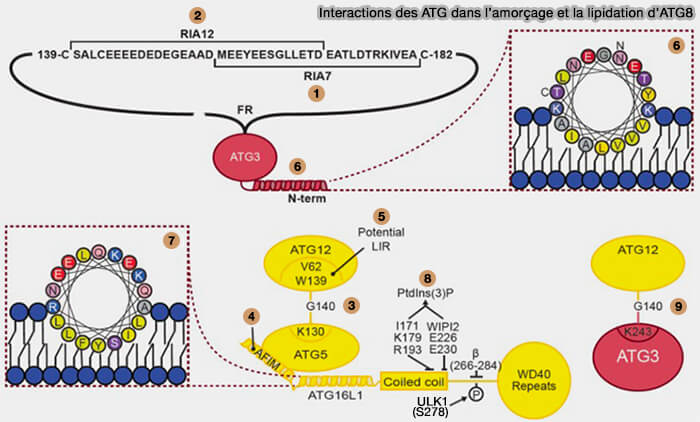
Récapitulation des interactions
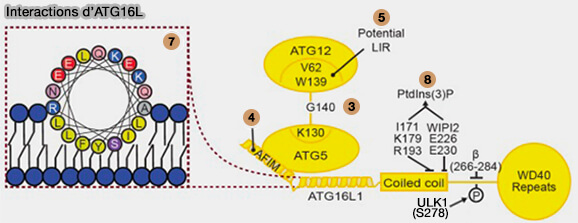
a. ATG3 interagit avec ATG7 et ATG12 par le biais de motifs situés dans la région flexible (FR), résidus 88-192, ces motifs sont appelés RIA7 (1) et RIA12 (2) respectivement.
G140 d'ATG12 est conjugué à K130 d'ATG5 (3) ou K243 d’ATG3 (9).
Le motif LIR, constitué par V62 et W139 d’ATG12 (5) fixerait le complexe à la membrane d’un autophagosome pendant la lipidation (5).
b. ATG16L1 interagit avec ATG5 par son hélice N-terminale, i.e. 13–28 (4).
c. ATG3, par son hélice amphipathique à l’extrémité N-terminale (aa 1-26), assure une liaison membranaire essentielle à la lipidation des protéines ATG8 (6).
Interactions des ATG dans l’amorçage et la lipidation d’ATG8
(Figure : vetopsy.fr d'après Lystag et Simonsenl)
d. L’interaction membranaire d'ATG16L1 est réalisée par de multiples interactions :
une hélice amphipathique près de l’extrémité N-terminale assure la liaison membranaire et est nécessaire à la lipidation de l’ATG8 (7),
les résidus conservés dans le domaine coled-coil,
son interaction avec WIPI2 (8) assurent la liaison à PI(3)P, la liaison membranaire à travers l’insert spécifique de l’isoforme β est importante pour la lipidation de la protéine ATG8 aux endosomes/lysosomes endommagés.
L’insert spécifique à l’isoforme β est une zone fortement modifiée post-traductionnellement dans ATG16L1 et comprend un site de phosphorylation confirmé pour ULK1 sur S278.





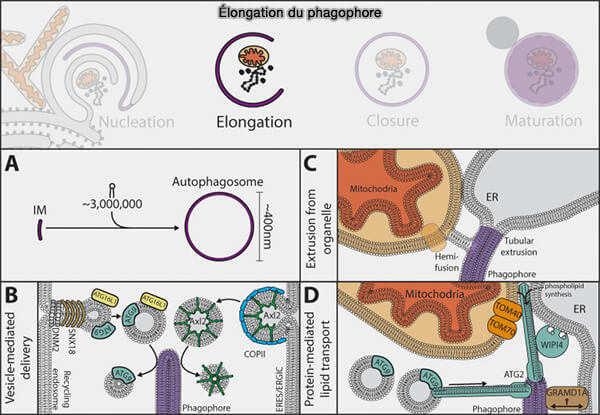
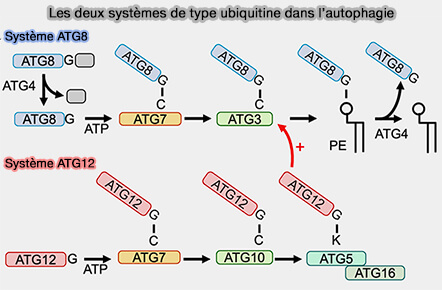
 tableau).
tableau).