


Mitochondries
Sites de contact membranaire (MCS ou MAM)
- En construction

Les sites de contact membranaire (MCS) sont des régions d'apposition étroite (≤ 30 nm) entre deux organites, établissant des microdomaines qui permettent un échange de matériel.
Les progrès récents de la biologie des MCS ont révélé leurs rôles clés dans les échanges inter-organites.
(Figure : vetopsy.fr d'après Inserm)
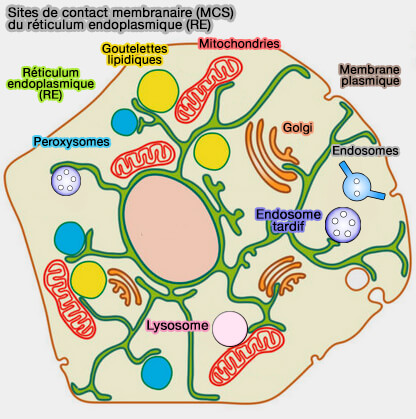
1. Le réticulum endoplasmique (RE) est l'organite qui comporte le plus grand nombre de sites de contact membranaire (MCS) avec de multiples organites.

Les MCS du réticulum endoplasmique sont étudiés dans des chapitres spéciaux.
2. Les mitochondries interagissent aux MCS avec :
- le réticulum endoplasmique (RE) ,
- les lysosomes,
- les peroxysomes, avec lesquels elle ont de nombreux points communs (The Peroxisome-Mitochondria Connection: How and Why? 2017).
Les MCS des peroxysomes sont étudiés dans des chapitres spéciaux.
Les sites membranaires associées aux mitochondries sont aussi appelées MAM, i.e. Mitochondrial Associated Membranes.

Vous pouvez lire : Here, There and Everywhere: The Importance of ER Membrane Contact Sites (2018) et The functional universe of membrane contact sites (2019).
MCS mitochondries/RE
Les mitochondries interagissent largement avec le réticulum endoplasmique (RE) dans les cellules vivantes (Visualizing Intracellular Organelle and Cytoskeletal Interactions at Nanoscale Resolution on Millisecond Timescales 2018).
Les MCS mitochondries/RE sont favorisées par l'interaction VAPB/PTPIP51 (Protein Tyrosine Phosphatase Interacting Protein 51), cette dernière possédant un motif FFAT (Phospholipid transfer function of PTPIP51 at mitochondria-associated ER membranes 2021).
Transport de molécules
Les MCS mitochondries/RE sont impliqués dans le transport de certaines molécules, sans que l'on sache comment les processus fonctionnent exactement.
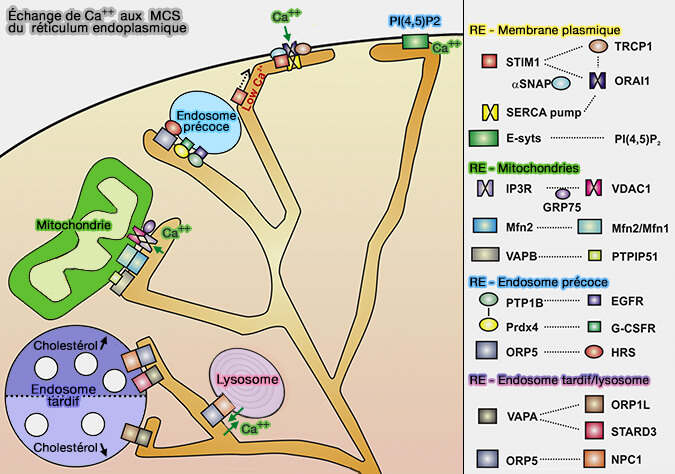
Calcium (Ca++)
1. Le calcium Ca++ peut être transporté de la lumière du RE vers la matrice mitochondriale.
- Le calcium est libéré du RE via le canal IP3R (Inositol 1,4,5-triphosphate Receptor) et acheminé vers le canal VDAC (Voltage Dependent Anion Channel) de la membrane mitochondriale externe (OMM).
- Après avoir traversé l'OMM, Ca++ est pris en charge par le canal MCU (Mitochondrial Calcium Uniporter) de la membrane mitochondriale interne (IMM) pour passer dans la matrice mitochondriale (Integrative genomics identifies MCU as an essential component of the mitochondrial calcium uniporter 2011).
2. Grp75 est un régulateur cytosolique du complexe IP3R-VDAC qui favorise l'interaction entre les canaux pour augmenter l'efficacité de l'absorption mitochondriale de Ca++ (Chaperone-mediated coupling of endoplasmic reticulum and mitochondrial Ca2+ channels 2006).
3. Deux protéines pourraient réguler ces contacts :
- la mitofusine 2 (Mfn2) de l'OMM, i.e. protéine qui participe aussi à la fusion mitochondriale,
- PDZD8, la protéine membranaire du RE qui favorise aussi la maturation des endosomes et les transfert de lipides (
 MCS à trois voies et ER-mitochondria tethering by PDZD8 regulates Ca2+ dynamics in mammalian neurons 2017).
MCS à trois voies et ER-mitochondria tethering by PDZD8 regulates Ca2+ dynamics in mammalian neurons 2017).
(Figure : vetopsy.fr d'après Burgoyne et coll)
Lipides
Les mitochondries et le réticulum endoplamique (RE) sont nécessaires à la biosynthèse des lipides, i.e. une navette devrait permettre le passage de molécules intermédiaires dans ce processus.
Vous pouvez lire : In situ architecture of the ER-mitochondria encounter structure (2023).
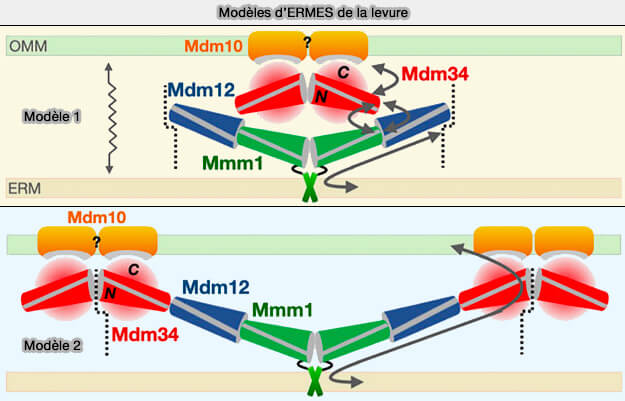
Complexe ERMES
1. Le complexe ERMES (ER-Mitochondria Encounter Structure) est formé de quatre sous-unités qui relie le RE aux mitochondries (Conserved SMP domains of the ERMES complex bind phospholipids and mediate tether assembly 2015).
ERMES n'est pas trouvé chez les métazoaires et semble principalement limité aux levures et aux champignons.
La structure, les différents composants et les modèles du complexe ERMES sont étudiés dans un chapitre spécial.
a. Trois sous-unités contiennent des domaines SMP (Synaptotagmin-like Mitochondrial lipid-binding Proteins), qui forment une longue cavité hydrophobe pour transférer les lipides entre les membranes.
b. Mdm10 est une porine mitochondriale
Remarque : la GTPase Gem1 liant le calcium, un orthologue des GTPases Miro, peut être considéré comme une cinquième sous-unité.
2. ERMES transfère la phosphatidylsérine (PS) et la phosphatidylcholine (PC) du RE à la membrane mitochondriale.
(Figure : vetopsy.fr d'après Montoro et coll)
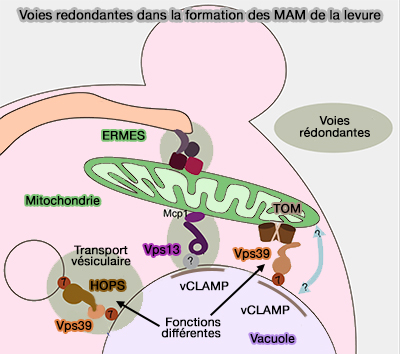
Toutefois, ERMES n'est pas essentiel car car il existe des voies redondantes, par exemple dans les vCLAMP (vacuolar and mitochondrial patch), i.e. sites de contact membranaire vacuole/mitochondrie (Vps39 Interacts with Tom40 to Establish One of Two Functionally Distinct Vacuole-Mitochondria Contact Sites 2018).
- Vps39, qui peut aussi être utilisé par le complexe HOPS avec Ypt7/Rab7, la petite GTPase Rab vaculaire, peut se lier avec Tom40, sous-unité principale du complexe TOM (Translocase of the Outer membrane), translocase de la membrane externe (OMM) pour former les vCLAMP.
- Vps13 peut aussi se lier avec Mcp1.
3. D'autres fonctions d'ERMES montrent son implication dans :
- l'importation et l'assemblage des protéines mitochondriales,
- la signalisation du calcium et la régulation de ses pools cellulaires,
- la distribution des mitochondries et de leur ADN,
- l'homéostasie du fer et la biosynthèse de la coenzyme Q .
(Figure : vetopsy.fr d'après Egea)
Lam6/Ltc1
Lam6/Ltc1, protéine du réticulum endoplasmique, appartenant aux LTP (Lipid Tranfer Protein) à domaine START-like, peut transférer les stérols par leurs interactions avec TOM70/71 de la membrane mitochondriale externe (OMM) (Ltc1 is an ER-localized sterol transporter and a component of ER–mitochondria and ER–vacuole contacts 2015 et Lam6 Regulates the Extent of Contacts between Organelles 2015).
Une autre protéine START-like, Lam4p pourrait jouer ce rôle entre le RE et la membrane plasmique (A new family of StART domain proteins at membrane contact sites has a role in ER-PM sterol transport 2015).
Tubulation et fission mitochondriale
1. Les mitochondries associées au RE sont localisées préférentiellement sur les microtubules acétylés (ER sliding dynamics and ER–mitochondrial contacts occur on acetylated microtubules 2010).
- Ces microtubules sont des voies privilégiées pour la kinésine-1 (Concerted action of kinesins KIF5B and KIF13B promotes efficient secretory vesicle transport to microtubule plus ends 2020).
- La kinésine 1 est un moteur à la fois pour le RE et les mitochondries.
Cette association permet la tubulation dynamique des mitochondries.
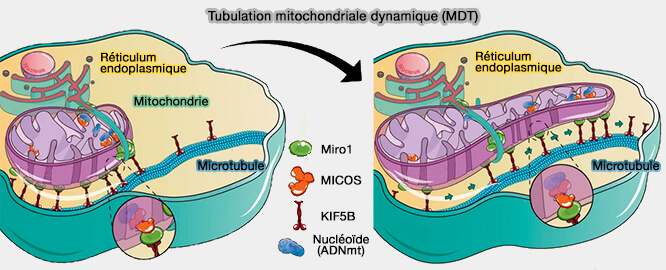
(Figure : vetopsy.fr d'après Elbaz-Alon et coll)
2. Les MCS mitochondries/RE jouent un rôle majeur dans la fission mitochondriale.
- Les tubules du réticulum endoplasmique s'enroulent autour des mitochondries en formant des MCS qui permettent de préciser l'endroit où la fission doit se produire.
- À ces contacts, les protéines mises en jeu dans la fission sont recrutés.
La fission mitochondriale est étudiée dans un chapitre spécial.
3. La constriction de l'IMM associée au RE se produit avant la constriction OMM (Constriction of the mitochondrial inner compartment is a priming event for mitochondrial division 2017).
Les nucléoïdes contenant l'ADNmt, aux sites de contact RE-mitochondries, interagissent avec le complexe protéique de la membrane interne MICOS (ER-mitochondria contacts promote mtDNA nucleoids active transportation via mitochondrial dynamic tubulation 2020).
- MICOS (MItochondrial contact site and Cristae Organizing System), est associé à la GTPase atypique de la membrane externe Miro1, qui est concentrée aux sites de contact RE-mitochondries et recrute les kinésines KIF5B sur les mitochondries.
- Lorsque la tubulation dynamique pilotée par KIF5B se produit, les nucléoïdes sont transportés via l'axe d'interaction nucléoïde/MICOS/Miro1/KIF5B.
(Figure : vetopsy.fr d'après Wu et coll)
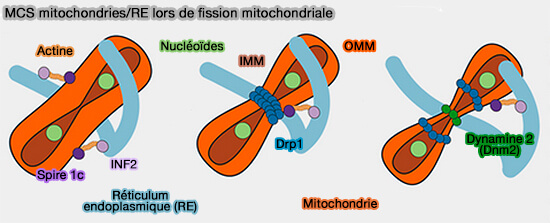
4. La constriction de l'OMM dépend aussi du RE.
La mitochondrie est d'abord, comprimée dans les MCS par des assemblages actine-myosine par :
- INF2, une formine inversée (INverted Formin 2) localisée dans le RE (), (An Actin-Dependent Step in Mitochondrial Fission Mediated by the ER-Associated Formin INF2 2013)
- Spire 1c, une protéine de nucléation d'actine mitochondriale (A mitochondria-anchored isoform of the actin-nucleating spire protein regulates mitochondrial division 2015).
Puis, Drp1, la dynamine de la division mitochondriale, est ciblée sur ses zones par des protéines adaptatrices, i.e. Fis1, Mff, MiD49 et MiD51 et s'oligomérise autour de l'OMM pour provoquer la constriction mitochondriale, achevé par la dynamine 2 ou Dnm2 ( ciblage des sites de fission).
Remarque : Il semblerait que les MCS mitochondries/RE contenant la mitofusine 2 (Mfn2) ne soient pas indispensables à la fusion mitochondriale ( autres rôles des mitofusines).
MCS mitochondries/lysosomes
1. Les mitochondries interagissent aussi avec les lysosomes et le réticulum endoplasmique (RE) dans les échanges de lipides.
Les échanges de lipides dans les MCS sont détaillés dans un chapitre spécial.
2. Les lysosomes en mouvement peuvent extraire de fins tubules des mitochondries.
Le rôle des MCS mitochondrie/lysosomes dans le trafic des lysosomes est détaillé dans un chapitre spécial.
Est-ce que les MCS à trois voies induites par PDZD8 entre le réticulum endoplasmique, les endosomes tardifs, et les mitochondries pourraient être impliquées dans les deux processus (dynamique du RE et endosomes tardifs/lysosomes) ?
Toutefois, cette interaction complexe a essentiellement immobilisé les organites (PDZD8 interacts with Protrudin and Rab7 at ER-late endosome membrane contact sites associated with mitochondria 2020).
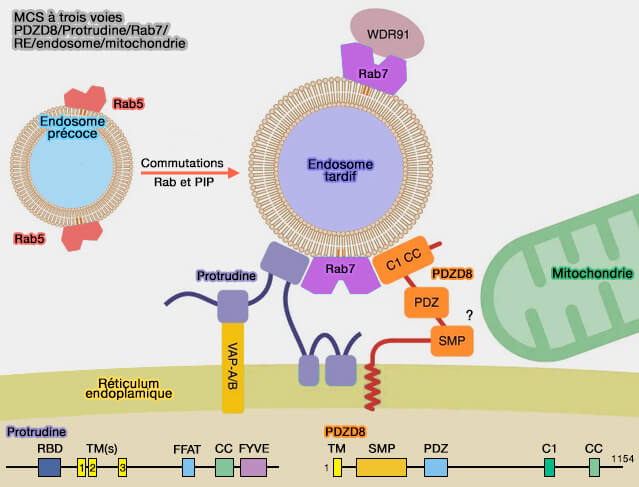
(Figure : vetopsy.fr d'après Elbaz-Alon et coll)
Biologie cellulaire et moléculaireMembrane plasmiqueNoyauCytoplasmeMitochondriesSystème endomembranaireRéticulum endoplasmiqueAppareil de GolgiEndosomesLysosomesPeroxysomesProtéasomesCytosqueletteMicrofilaments d'actineFilaments intermédiairesMicrotubulesMatrice extracellulaireReproduction cellulaireBiochimieTransport membranaireMoteurs moléculairesVoies de signalisation