


Système endomembranaire
Complexes TRAPP

Les complexes TRAPP (TRAnsport Protein Particle) sont des complexes à nombreuses sous-unités, impliqués dans le trafic membranaire, i.e. sécrétion et autophagie, aussi bien chez la levure que chez les métazoaires.
TRAPP est un complexe protéique modulaire à nombreuses sous-unités hautement conservées qui agit comme (The two TRAPP complexes of metazoans have distinct roles and act on different Rab GTPases 2018) :
- protéine de transport entre le réticulum endoplasmique (RE) et l'appareil de Golgi,
- GEF (Guanine nucleotide exchange factor) pour les petites GTPases Rab/Ypt.
Complexes TRAPP
La famille TRAPP est une famille de complexes hautement conservée de la levure aux métazoaires.
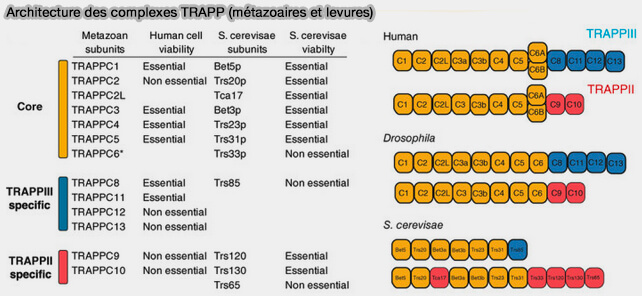
(Figure : vetopsy.fr d'après Galindo et Munro)
1. Chaque complexe est composé d’un noyau identique de protéines, i.e. TRAPP-I, i.e. composé de deux sous-complexes, TRAPPC1-TRAPPC3-TRAPPC4-TRAPPC6 et TRAPPC1-TRAPPC3-TRAPPC5 (The Architecture of the Multisubunit TRAPP I Complex Suggests a Model for Vesicle Tethering 2006).
- Chez la levure, les homologues sont Bet5p, Trs20p, Bet3p, Trs23p, Trs31p, Trs33p, i.e. TrsN, N étant la taille en kDa.
- Trs20p/Sedlin serait un adaptateur nécessaire à l’association du noyau TRAPP avec des sous-unités plus grandes pour former TRAPP II et TRAPP III.
- On y ajoute le plus souvent Tca17/TRAPPC2L, un paralogue de Trs20.
Remarque : TRAPPC3 est présent deux fois.

Vous pouvez lire pour une analyse des sous-unités et une bibliographie récente : TRAPPopathies: Severe Multisystem Disorders Caused by Variants in Genes of the Transport Protein Particle (TRAPP) Complexes (2024).
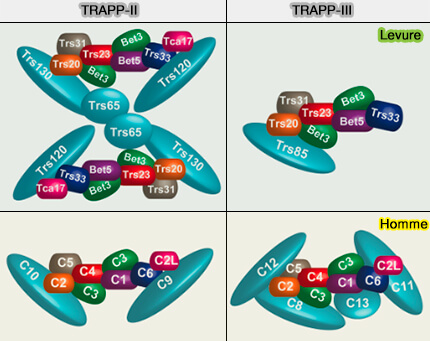
(Figure : vetopsy.fr d'après Brunet et Sacher)
2. Les deux complexes TRAPP-II et TRAPP-III comprennent des protéines supplémentaires, spécifiques au complexe, qui définissent leur identité, leur localisation et leur fonction (In Sickness and in Health: The Role of TRAPP and Associated Proteins in Disease 2014).
a. À ce jour, onze sous-unités de TRAPP conservées entre la levure et l’homme ont été identifiées (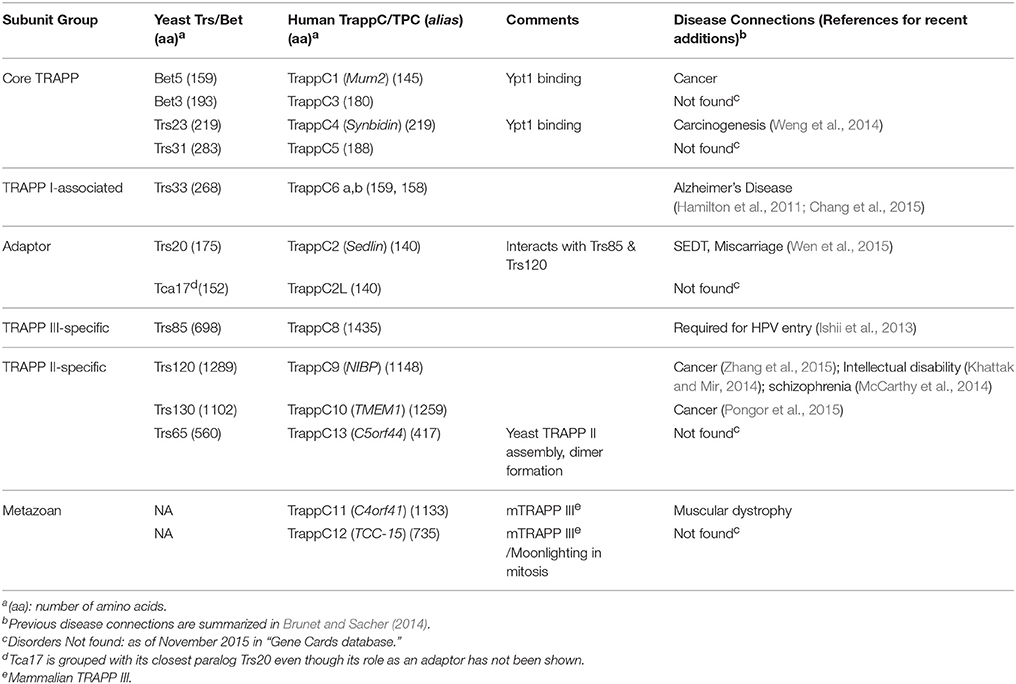 tableau).
tableau).
- Sept sous-unités sont petites, i.e. 140 à 283 acides aminés.
- Les autres sont plus grandes, i.e. 417 à 1435 résidus, et sont exclusives aux complexes TRAPP II ou TRAPP III.
b. Deux sous-unités supplémentaires, TRAPPC11/12, sont spécifiques aux métazoaires.
Remarque : cette liste n'est pas figée et on peut rajouter TRAPPC13, et de plus, on ne sait pas s'il existe d'autres sous-unités ou si certaines ne sont que des adaptateurs pour les complexes comme Trs20 ou Trs85 (voir pour le détail dans TRAPP Complexes in Secretion and Autophagy 2016).
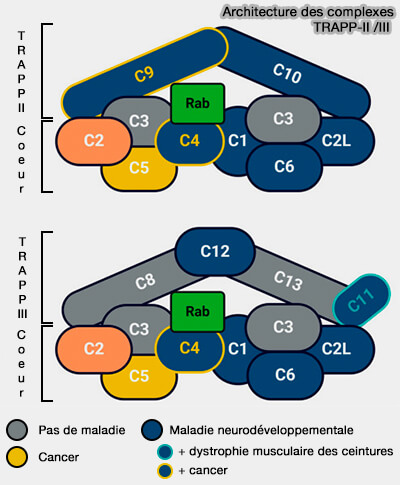
(Figure : vetopsy.fr d'après Hall et coll)
3. Alors que la structure moléculaire du complexe TRAPP central est résolue, l’architecture des plus grands complexes TRAPP, y compris leur existence en tant que dimères et multimères, est moins claire.

Compte tenu de la taille globale des complexes TRAPP et du nombre de sous-unités, leurs protéines ont des partenaires d'interaction uniques, permettant ainsi aux complexes de coordonner des événements distincts même dans une voie commune.
4. Les complexes TRAPP sont impliqués dans des maladies chez l'homme, appelées TRAPPopathies (TRAPPopathies: Severe Multisystem Disorders Caused by Variants in Genes of the Transport Protein Particle (TRAPP) Complexes 2024).
Rôles dans le trafic vésiculaire
Nous ne développerons pas ce chapitre pour l'instant.
Vous pouvez lire pour une analyse et une bibliographie récente : The TRAPP complexes: oligomeric exchange factors that activate the small GTPases Rab1 and Rab11 (2022).
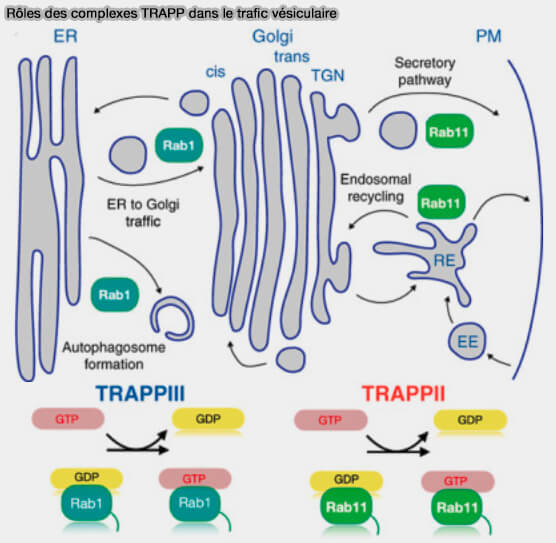
(Figure : vetopsy.fr d'après Galindo et Munro)
Rôles de TRAPP-III dans l'autophagie
Le rôle de TRAPP-III est encore obscur dans l'autophagie, bien que plusieurs études récentes aient montré un rôle pour TRAPP-III dans ce processus (TBC1D14 regulates autophagy via the TRAPP complex and ATG9 traffic 2015 et TRAPPC13 modulates autophagy and the response to Golgi stress 2017).
Les mutations de TRAPPC11, TRAPPC12 et TRAPPC8 affectent l'autophagie.
TRAPPC8
1. Chez la levure, Trapp-III interagit avec les vésicules contenant de l'Atg9 (Atg9 Vesicles Recruit Vesicle-tethering Proteins Trs85 and Ypt1 to the Autophagosome Formation Site 2012).
- Cette interaction implique TRS85, l'homologue de TRAPPC8.
- Le recrutement initial de Trapp-III sur la vacuole est dépendant de l'ATG9.
2. Chez les mammifères, il semble que TRAPPC8 fonctionne également en amont de la formation du phagophore (TBC1D14 regulates autophagy via the TRAPP complex and ATG9 traffic 2015).
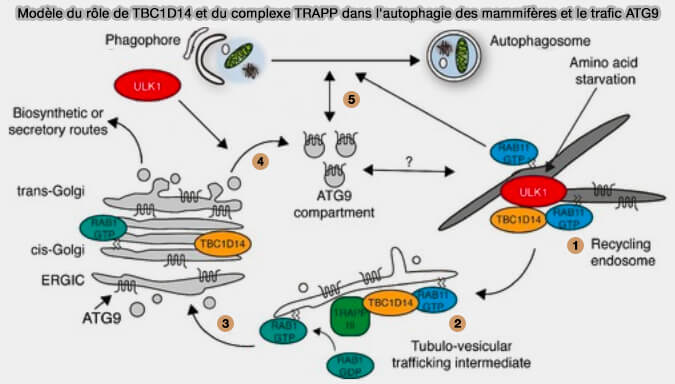
(Figure : vetopsy.fr d'après Lamb et coll)
Les endosomes de recyclage abritent une population de TBC1D14, régulateur négatif de l'autophagie, liée à RAB11, et une population de molécules ATG9 qui peuvent circuler vers et depuis le compartiment ATG9.
- 1-2 Lors d’une privation d’acides aminés, TBC1D14 et RAB11 induisent une étape de trafic vésiculaire des membranes positives à RAB11 vers celles positives à RAB1 au niveau d’un intermédiaire de transport tubulo-vésiculaire (TBC1D14 regulates autophagosome formation via Rab11- and ULK1-positive recycling endosomes 2012).
- 3. Cela se traduit par le recyclage d’ATG9 vers des compartiments de Golgi régulés par RAB1.
Chez la levure, l'activation du complexe ULK et la translocation du réticulum endoplasmique (RE) seraient dépendantes de la liaison à l'effecteur C9orf72 de Rab1 (The C9orf72 protein interacts with Rab1a and the ULK1 complex to regulate initiation of autophagy 2016).
- 4. ATG9 peut être acheminé vers le compartiment ATG9 de manière dépendante d’ULK1.
- 5. Cela maintient le compartiment ATG9 lors de dénutrition, ce qui peut alors à la fois contribuer et favoriser la formation d’autophagosomes à partir d’autres sources membranaires, y compris à partir d’endosomes de recyclage positif RAB11.
TRAPPC11
1. Le recrutement d'ATG2B et de WIPI4 au phagophore dépend de TRAPPC11 et implique son extrémité C-terminale (TRAPPC11 functions in autophagy by recruiting ATG2B- WIPI4/WDR45 to preautophagosomal membranes 2019).
WIPI4 semble aussi réguler l'association de l'ATG9A avec les membranes LC3.
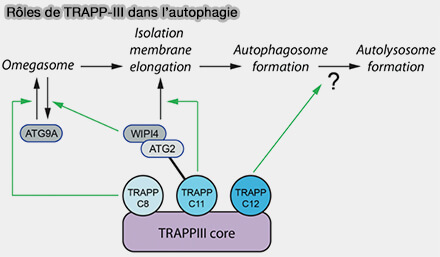
(Figure : vetopsy.fr d'après Stanga et coll)
Comme l'ATG9A ne s'associe que transitoirement aux membranes LC3, on peut penser que le recrutement dépendant de TRAPPC11 d'ATG2B-WIPI4 est la libération d'ATG9A à partir des phagophores/omégasomes (Dynamic and transient interactions of Atg9 with autophagosomes, but not membrane integration, are required for autophagy 2012).
2. Il participerait ainsi à la fermeture de l'autophagosome.
La déplétion de TRAPC11 se traduit par un phénotype similaire à celui des KO de l'ATG2A/B et de l'ATG2A-mLIR.
TRAPPC12
Le rôle de l'étape de l'action de TRAPPC12 est obscur, mais il survient après TRAPPC11, donc après la fermeture du phagosome.
- TRAPPC12 pourrait recruter un facteur tardif nécessaire pour la fusion, comme le partenaire TECPR1, une protéine précédemment impliquée dans la fusion autophagosome/lysosome.
- Il pourrait faciliter l'activité d'une protéine de membrane autophagosomique dentifiée nécessaire à la formation d'autolysosomes.
Retour à l'autophagie
Biologie cellulaire et moléculaireConstituants de la celluleSystème endomembranaireRéticulum endoplasmiqueAppareil de GolgiEndosomesLysosomesPeroxysomesAutophagieMacroautophagieMicroautophagieAutophagie chaperonnes (CMA)Autophagie non canoniqueProtéines ATGSystème UPS (Ubiquitine-Protéasome)Transport membranaireTrafic vésiculaireFusion/fission membranaireEndocytoseVoie sécrétoireMoteurs moléculairesVoies de signalisation