Système endo-lysosomal
Réticulum endoplasmique
Sites de contact membranaire (MCS) du RE
Endosomes multivésiculaires (MVE/MVB) et lysosomes
2. Trafic, positionnement, fission et fusion
Les sites de contact membranaire (MCS) du réticulum endoplasmique avec les endosomes multivésiculaires (MVE/MVB) et les lysosomes sont impliquées dans le trafic et le positionnement des endosomes/lysosomes, la fission et la fusion membranaires.
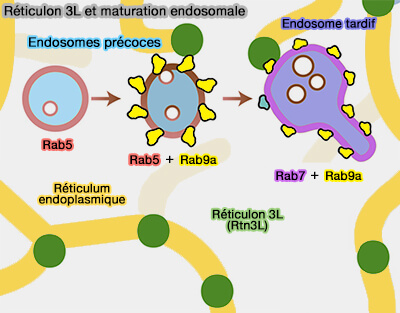
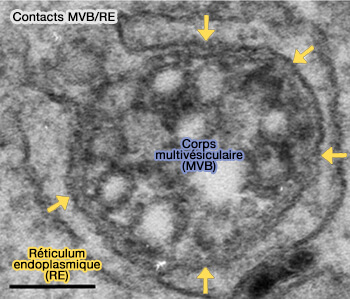
Contacts MVB/RE
(Figure : vetopsy.fr d'après Wong et coll)
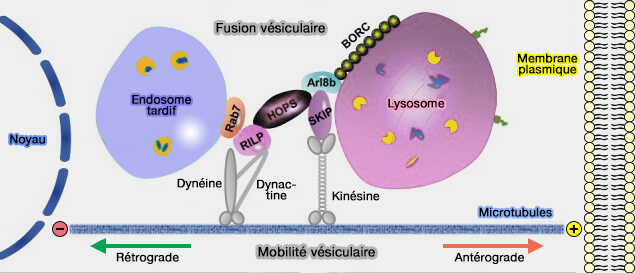
Modèle de transport des endosomes
(Figure : vetopsy.fr d'après Khattern et coll)
Remarque : le déplacement des organites peut aussi provoquer la croissance des microtubules est appelé " auto-stop " (hitchhiking en anglais).
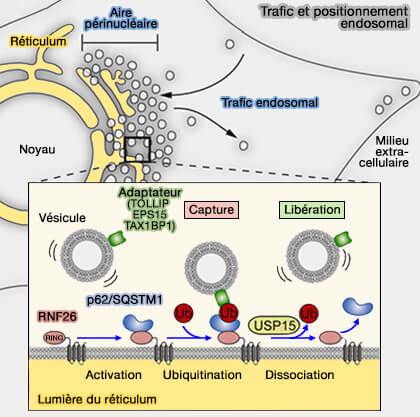
Positionnement des endosomes par RNF26
Les organites endo-lysosomaux, bien qu'ils puissent se déplacer de manière bidirectionnelle, sont localisés, à l'état d'équilibre, dans la région périnucléaire appelée MTOC (MicroTubule-Organizing Center).
Trafic et positionnement endosomal
(Figure : vetopsy.fr d'après Jongsma et Coll)
L'ubiquitine entre en contact avec la protéine adaptatrice p62/SQSTM1, interagit à son tour avec diverses protéines adaptatrices endo-lysosomales, notamment ( devenir des protéines ubiquitinées) :
La structure de ces protéines adaptatrices comprend des domaines spécifiques de liaison à l'ubiquitine et à la membrane qui permettent d'attacher leurs compartiments respectifs au RE périnucléaire sur lequel est localisé RNF26.
1. Ce mécanisme implique les protéines VAP, i.e. VAPA et son paralogue VAPB, protéines endoplasmiques qui participent aussi au contrôle de la liaison des kinésines et de la dynéine aux lysosomes.
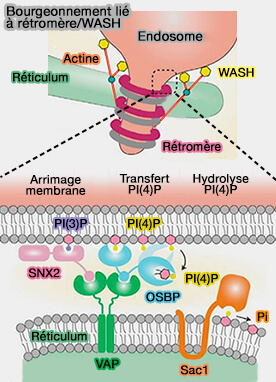
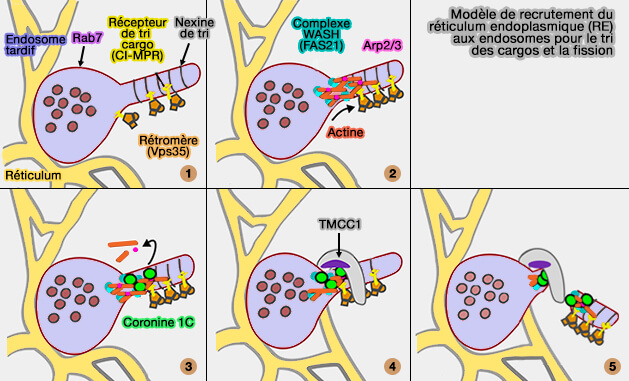
Ces interactions conduisent à la nucléation de l'actine dépendante de PI(4)P et du complexe WASH, un activateur de la protéine de nucléation Arp2/3, pour activer le bourgeonnement des tubules contenant des rétromères.
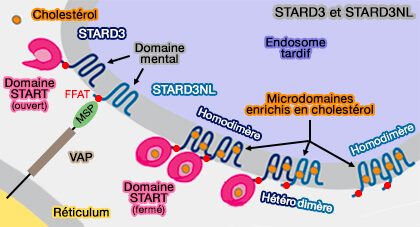
STARD3 et STARD3NL
(Figure : vetopsy.fr d'après Wilhelm)
Isoformes de la spastine et microtubules
(Figure : vetopsy.fr d'après Allison et coll)
1. L'endosome tardif bourgeonne, le cargo attaché au rétromère et les nexines de tri stabilisant l'élongation.
La perturbation de cette interaction a conduit à un mauvais tri des enzymes lysosomales et à des anomalies lysosomales, ce qui est probablement la raison sous-jacente pour laquelle les mutations de la spastine provoquent une paraplégie spastique héréditaire.
Ces exemples illustrent comment le transport endolysosomal est intégré à la fission et à la fusion, et mettent en évidence le rôle du RE dans la régulation de ces processus.
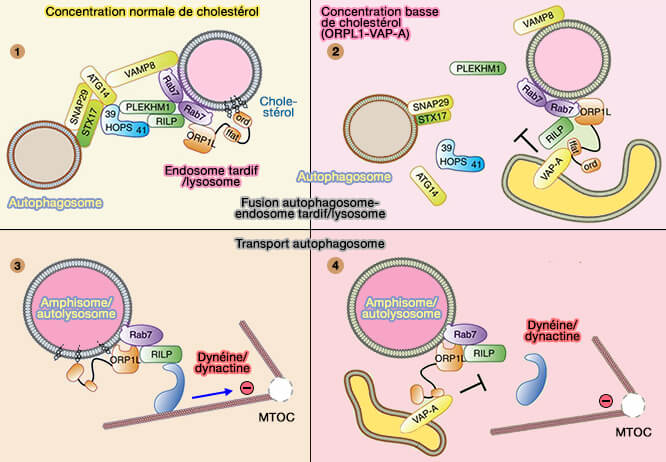
Modèle de ORPL1 pour la fusion et le trafic
(Figure : vetopsy.fr d'après Wijdeven et coll)





 hitchhiking en anglais).
hitchhiking en anglais).