


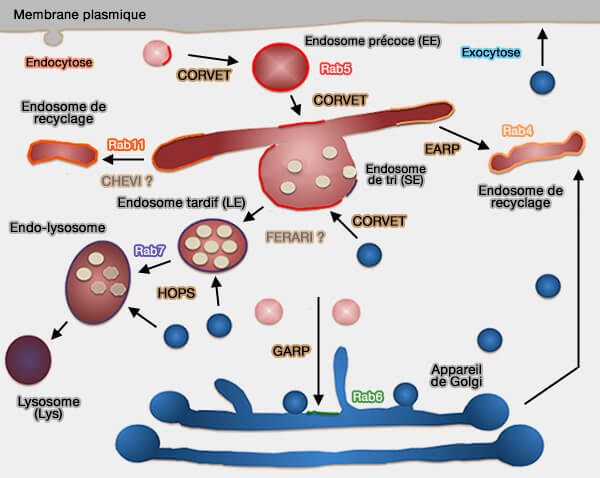
Système endo-lysosomal
Endosomes tardifs
Rab7

La maturation des endosomes précoces en endosomes tradifs nécessite la petite GTPase Rab7 et des lipides spécifiques, en particulier PI(3,5)P2 et LBPA (ou BMP).
(Figure : vetopsy.fr d'après Spang)
Les endosomes tardifs ont généralement un pH inférieur à celui des endosomes précoces, de l'ordre de 5,5 à 6,0, grâce à la fonction d'une pompe à protons V-ATPase ( pH des différents compartiments subcellulaires).
pH des différents compartiments subcellulaires).
La maturation des endosomes précoces et la fusion des endosomes tardifs nécessitent :
- des lipides spécifiques, en particulier PI(3,5)P2 et LBPA/BMP ( phospholipides et endosomes),
- la petite GTPase Rab7 (Ypt7 de la levure),
-
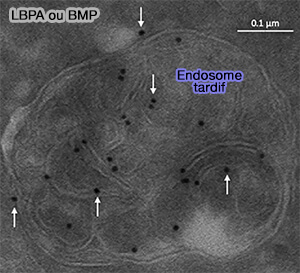
LBPA (ou BMP)
(Figure : vetopsy.fr d'après Gruenberg) - les protéines SNARE.
Rab7
Vue d'ensemble
1. Les endosomes tardifs contiennent Rab7 (Ypt7 de la levure) à la place de Rab5 (Vps21 de la levure) des endosomes précoces, i.e. RAb5 est remplacé par Rab7 en seulement quelques minutes (Identification of the Switch in Early-to-Late Endosome Transition 2010).
Rab7 est également présente sur les endosomes précoces et aide à recruter le rétromère (Rab GTPase regulation of retromer-mediated cargo export during endosome maturation 2012).
2. Rab7 joue un rôle majeur, grâce au complexe HOPS et aux protéines SNARE, dans la fusion ( fusion endo-lysosomale) :
- des endosomes tardifs (Rab7 Regulates Late Endocytic Trafficking Downstream of Multivesicular Body Biogenesis and Cargo Sequestration 2009 et Rab Proteins and the Compartmentalization of the Endosomal System 2014),
- des endosomes tardifs avec les lysosomes, en formant un compartiment intermédiaire, l'endo-lysosome.

Remarque : on trouve aussi la petite GTPase Rab7a et son GAP TBC1D5 ( petites GTPases et rétromère) sur :
- les endosomes de tri EEA1+,
- le réseau trans-golgi (TGN),
- le réticulum endoplasmique (RE),
- les mitochondries,
- les gouttelettes lipidiques (Lipid Droplets ou LD).
Conversion de Rab5 à Rab7
La maturation endosomale, selon un modèle séduisant, implique la conversion séquentielle à travers le cycle GTPase d'un Rab pour le suivant ( Rab et maturation endosomale).
- Le premier Rab recrutant un GEF (Guanine nucleotide Exchange Factor), i.e. stimulant la libération de guanosine diphosphate (GDP) pour permettre la liaison de la guanosine triphosphate (GTP), i.e. stimulant l'action pour le Rab suivant.
- Ce dernier pourrait à son tour recruter un GAP (GTPase-Activating Protein), i.e. accélérant l'hydrolyse du GTP en GDP, qui met fin à l'action du Rab précédent.
Activation de Rab7
(Figure : vetopsy.fr d'après Naslavsky et Caplan)
1. Les protéines SAND-1 ou Mon1a/b chez les vertébrés sont recrutées par Rab5-GTP des endosomes précoces et jouent un rôle essentiel dans ce remplacement par deux processus (Function of the Mon1-Ccz1 complex on endosomes 2014).
a. D'abord, Mon1, qui contient un domaine longin :
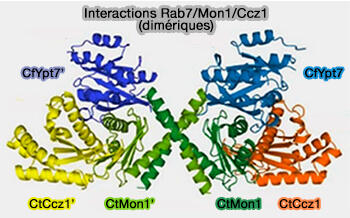
- recrute CCZ-1, la protéine de liaison de Rab7 sur les endosomes précoces, i.e. le complexe Rab5/Mon1/CCZ-1 sert de GEF à Rab7-GDP, i.e. étape nécessaire à l'activation de Rab7 (Identification of two evolutionarily conserved genes regulating processing of engulfed apoptotic cells 2010 et Architecture and mechanism of the late endosomal Rab7-like Ypt7 guanine nucleotide exchange factor complex Mon1–Ccz1 2017),
- interagit directement avec des protéines du complexe HOPS.
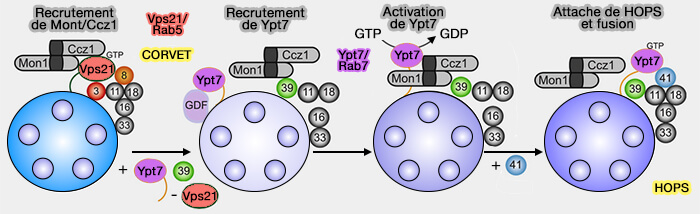
(Figure : vetopsy.fr d'après Nordmann et coll)
b. Mon1 inactive Rabex-5, la GEF (Guanine nucleotide exchange factor) qui recrute et maintient Rab5 sur la membrane des endosomes précoces (régulateurs de Rab5). De ce fait, elle met fin à la boucle d'activation positive et élimine Rab5 de la membrane (Identification of the Switch in Early-to-Late Endosome Transition 2010).
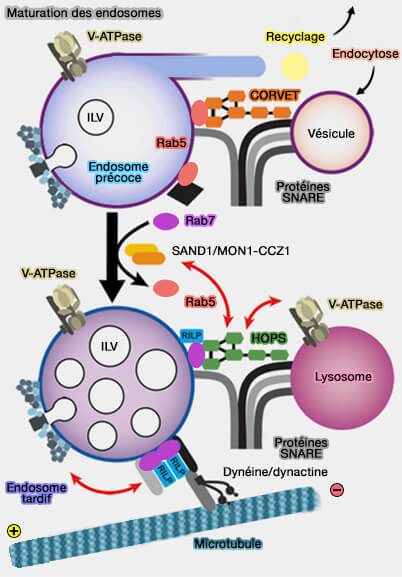
(Figure : vetopsy.fr d'après Solinger et Span)
2. L'activité du complexe Mon1/Ccz1, GEF pour Rab7, est fortement stimulée sur les membranes qui comprennent du PI(3)P et qui soutiennent également l'association membranaire du complexe GEF (The Mon1–Ccz1 GEF activates the Rab7 GTPase Ypt7 via a longin-fold–Rab interface and association with PI3P-positive membranes 2014).

La régulation spatiotemporelle de l’activité de SAND-1/Mon1 n’est pas encore bien comprise.
Remarque 1 : la GEF Mon1/Ccz1 est nécessaire et suffisante pour stabiliser Ypt7/Rab7 prénylé sur les membranes et, par la liaison du complexe HOPS à Ypt7-GTP, pour provoquer ensuite la fusion médiée par SNARE (A guanine nucleotide exchange factor (GEF) limits Rab GTPase–driven membrane fusion 2018).
Remarque 2 : Mont1 et Ccz1 possèdent un domaine longin présent aussi chez certaines protéines SNARE.
Rétroactions négatives
1. La GAP (GTPase-Activating Protein ou GTPase-Accelerating Protein) ARMUS/TBC-2 est impliquée dans l'inactivation de Rab5 pour promouvoir la conversion de Rab5 en Rab7 (Vps34 and the Armus/TBC-2 Rab GAPs: Putting the brakes on the endosomal Rab5 and Rab7 GTPases 2017 et Vps34 regulates Rab7 and late endocytic trafficking through recruitment of the GTPase-activating protein Armus 2016 et The VPS34 PI3K negatively regulates RAB-5 during endosome maturation 2017).
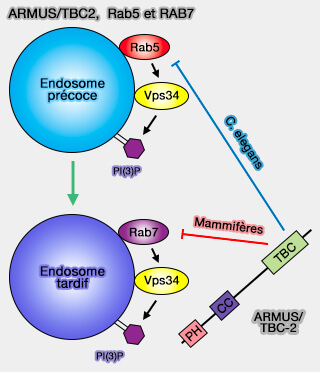
(Figure : vetopsy.fr d'après Law et coll)
La perte de Vps34, composant de deux complexes principaux (PI3KC3-CI et PI3KC3-C2) qui interviennent dans les différentes fonctions du trafic intracellulaire, entraîne une hypertrophie des endosomes précoces, par une augmentation de la concentration de PI(3)P qui provoque une fusion excessive.
Ce phénomène est semblable à celui observé par l'hyperactivité de Rab5, ce qui suggère une boucle de rétroaction négative pour l'inactivation des Rab5 et Rab7 (Vps34 regulates Rab7 and late endocytic trafficking through recruitment of the GTPase-activating protein Armus 2016 et The VPS34 PI3K negatively regulates RAB-5 during endosome maturation 2017).
La GAP ARMUS/TBC-2, localisée dans les endosomes tardifs, est caractérisée par :
- un domaine PH N-terminal, qui, contrairement à d'autres, peut se lier à PI(3)P et PI(4)P,
- un domaine coiled-coil (superhélice ou CC), qui interagit avec les protéines et/ou les lipides des membranes endosomales,
- un domaine C-terminal Tre-2/Bub2/Cdc16 (TBC)/Rab GAP.
ARMUS pourrait réguler l'activité de Rab7 ou, dans le cas de TBC-2 chez C. elegans, inactiver Rab5 pour favoriser la conversion de Rab5 en Rab7 pendant la maturation des endosomes (Vps34 regulates Rab7 and late endocytic trafficking through recruitment of the GTPase-activating protein Armus 2016 et The VPS34 PI3K negatively regulates RAB-5 during endosome maturation 2017).
2. La concentration en PI(3)P diminue aussi par l'interaction de WDR91/WDR81, SORF-1-SORF-2 de la levure avec le complexe Vps3 (Negative regulation of phosphatidylinositol 3-phosphate levels in early-to-late endosome conversion 2016).
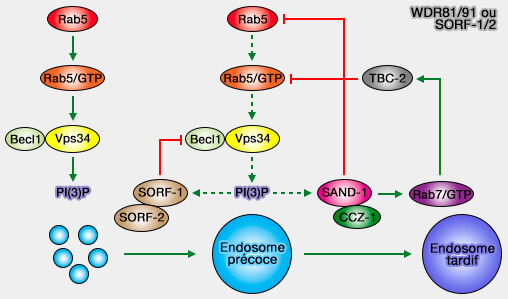
(Figure : vetopsy.fr d'après Liu et coll)
La perte de WDR91/WDR81, comme celle de Vps34 plus haut, entraîne une hypertrophie des endosomes.
- SORF-1-SORF-2 agissent au sein d'un complexe avec Beclin1 et leur perte augmente l'activité des PI3K.
- WDR91 sert d'effecteur Rab7 pour coupler le commutateur Rab endosomal avec la régulation à la baisse de Pi(3)P lors de la maturation de l'endosome (WDR91 is a Rab7 effector required for neuronal development 2017).
Complexe HOPS
Biologie cellulaire et moléculaireMembrane plasmiqueNoyauCytoplasmeMitochondriesSystème endomembranaireRéticulum endoplasmiqueAppareil de GolgiEndosomesLysosomesPeroxysomesProtéasomesCytosquelettemicrofilaments d'actineFilaments intermédiairesMicrotubulesMatrice extracellulaireReproduction cellulaireBiochimieTransport membranaire Moteurs moléculairesVoies de signalisation