Les peroxysomes sont de petits organites limités par une membrane qui se trouvent à l’intérieur des cellules.
Les peroxysomes participent à la respiration cellulaire, comme les mitochondries, i.e. ils consomment de l'oxygène en employant des enzymes oxydatives.
Leur nom provient de la leur capacité à neutraliser le peroxyde d'hydrogène (H202) très toxique, pour produire de l’eau.
Machineries moléculaires et protéines des membranes peroxysomales des mammifères
(Figure : vetopsy.fr d'après Islinger et coll)
Chez la levure, Inp1 et Inp2 (inheritance of peroxisomes 1 et 2) sont impliqués dans la transmission et la motilité des peroxysomes, Inp2 étant le récepteur membranaire de la myosine de type V Myo2 sur les peroxysomes, qui les entraîne le long des filaments d'actine. La petite GTPase Rho1 se lie à Pex25 et est impliquée dans le recrutement d'actine dans les peroxysomes.
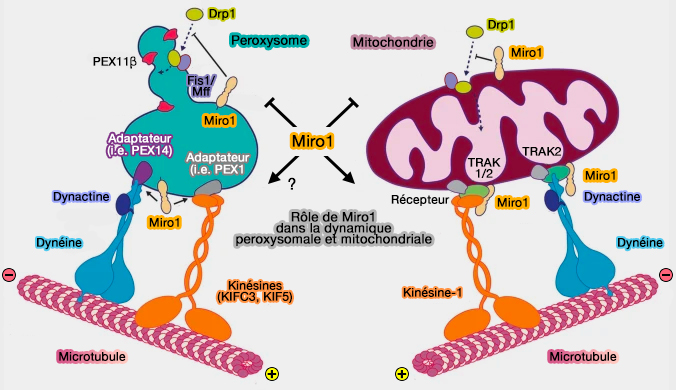
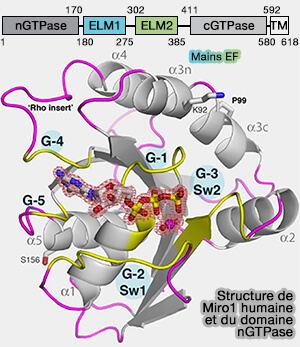
Rôle de Miro1 dans la dynamique peroxysomale et mitochondriale
(Figure : vetopsy.fr d'après Covill-Cooke et coll)
Les forces de traction médiées par Miro1 contribuent également à l'allongement de la membrane des peroxysomes et à leur prolifération et permettent désormais de relier le cytosquelette des microtubules et les forces de traction motrices à la formation de peroxysomes par croissance et division dans les cellules de mammifères (Mind the organelle gap - Peroxisome contact sites in disease 2018).
3. Toutefois, comme l'allongement et la division des peroxysomes peuvent se produire en l'absence de microtubules, des mécanismes indépendants mais coopératifs doivent exister.
Dans ce cas, MFF (Mitochondrial Fission Factor) pourrait agir comme un capteur de force lié à la membrane pour recruter la machinerie de fission sur des sites mitochondriaux soumis à des contraintes mécaniques.
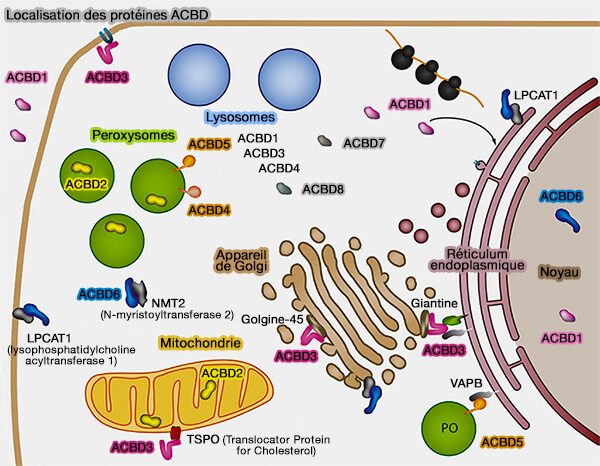
Remarque : ACBD2 se trouve dans la matrice peroxysomale.
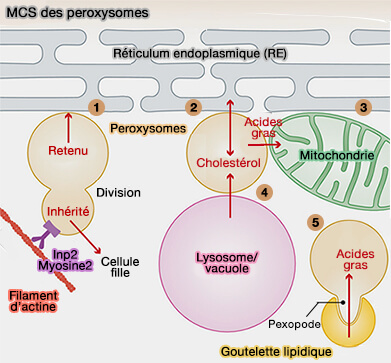
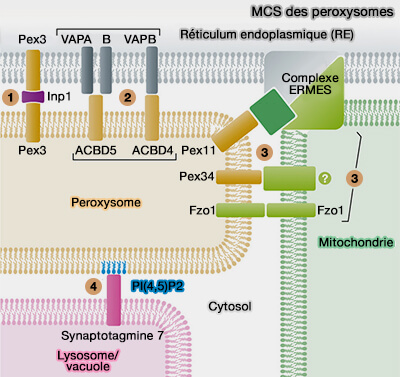
Sites de contact membranaire (MCS) des peroxysomes Figure : vetopsy.fr d'après Farré et coll)
3. Les fonctions de ces MCS sont nombreuses.
a. Les contacts peroxysomes-RE jouent un rôle dans le transfert des lipides. L'expansion et la croissance de la membrane peroxysomale est une condition préalable à la division et à la prolifération des peroxysomes ( biogenèse des peroxysomes).
Les défauts de fission des peroxysomes (par exemple en raison de la perte de la fonction Mff ou Drp1) entraînent des peroxysomes très allongés, suggérant un transfert constant de lipides du RE vers les peroxysomes.
Sites de contact membranaire (MCS) des peroxysomes Figure : vetopsy.fr d'après Farré et coll)Fzo1-Fzo1 (mitofusine de la levure), impliquées dans plusieurs MCS des mitochondries, des RE, des mélanosomes et les gouttelettes lipidiques (Lipid Droplets ou LD). Fzo1 interagit physiquement avec Pex19 et Pex14, ce qui suggère que Fzo1 pourrait être l'attache dans les deux organites.
Pex34 (PEX16 de mammifère) avec un partenaire mitochondrial indéfini (?) qui serait Fzo1 chez la levure.
Chez S. cerevisiae, les peroxysomes peuvent être localisés à côté d'une niche mitochondriale spécifique près du site de contact RE/mitochondrie, à proximité de l'endroit où se trouve le complexe pyruvate déshydrogénase (PDH), i.e. qui entre dans la biosynthèse de l'acétyl-CoA à partir du pyruvate, dans la matrice mitochondriale, suggérant ainsi une voie à trois voies.
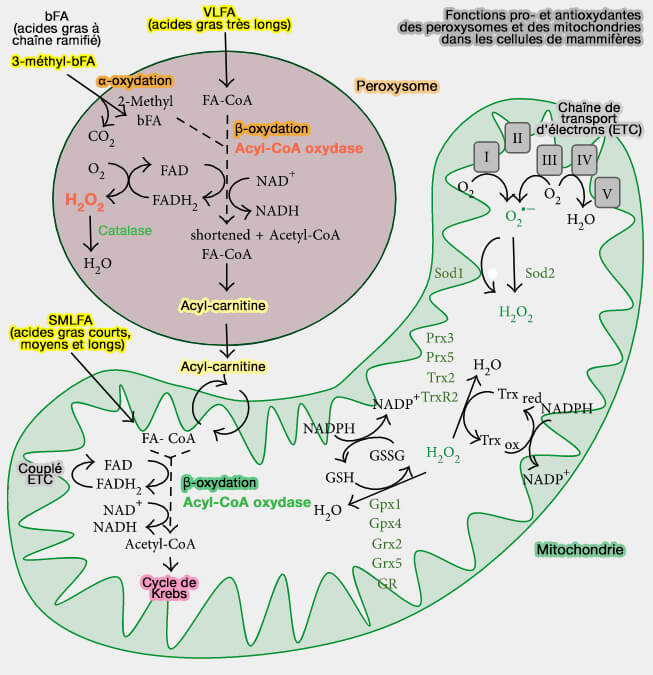
Fonctions pro- et antioxydantes des peroxysomes et des mitochondries (mammifères)
(Figure : vetopsy.fr d'après Pascual-Ahuir et coll)
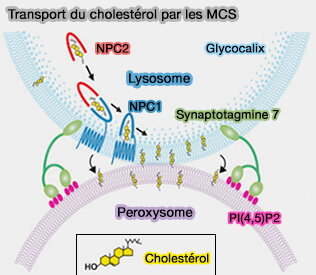
2. Les mutations dans NPC1 et NPC2, qui, ensemble, assurent le transport du cholestérol libre de la lumière vers la membrane limitante des lysosomes, provoquent une accumulation de cholestérol dans les lysosomes. Ces mutations provoquent la maladie de Niemann-Pick de type C (NPC), i.e. maladie neurodégénérative mortelle.
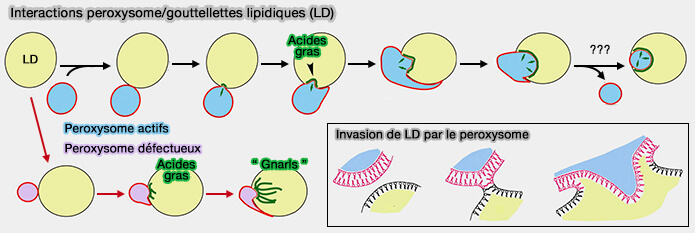
1. Les peroxysomes se lient aux LD et stimulent la dégradation des triglycérides, libérant des acides gras qui sont importés des LD et oxydés par les peroxysomes.
2. Lors des contacts, des saillies peroxysomales appelées pexopodes s'étendent dans le noyau des LD.
Ces saillies pourraient représenter des lieux d'hémi-fusion entre le feuillet externe de la membrane peroxysomale avec la monocouche phospholipidique des LD, tandis que le feuillet interne envahit le noyau LD.
Les pexopodes sont enrichis en protéines impliquées dans la β-oxydation, ce qui indique qu'ils pourraient être des endroits où les acides gras sont transférés des LD aux peroxysomes.
Remarque 1 : les peroxysomes qui sont incapables d'effectuer une β-oxydation provoquent l'accumulation d'acides gras, ce qui entraîne des " gnarls " (protubérances ?).




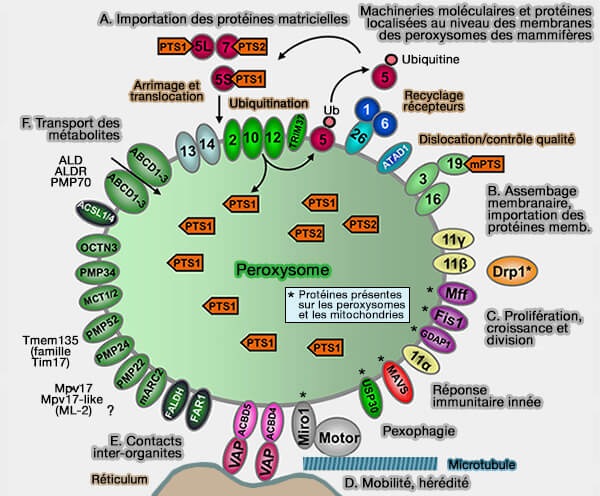

 tableau des protéines motrices impliquées).
tableau des protéines motrices impliquées).