Les propriétés des myosines, retrouvées dans les cellules musculaires et non-musculaires et dont il existe de nombreuses classes et de nombreuses isoformes, sont :
de se fixer sur l'actine-F, ou plutôt les actines dont il existe 6 isoformes chez le mammifères :
4 dans les muscles (αskeletal-actine, αcardiac-actine, αsmooth-actine, γcyto-actine),
2 cellulaires ubiquitaires (βcyto-actine et γcyto-actine).
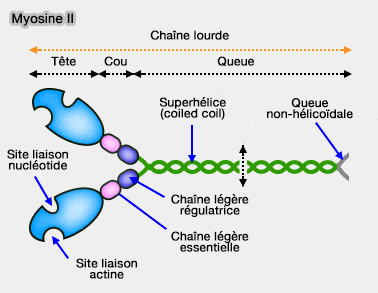
Structure de la myosine II
(Figure : vetopsy.fr)
d’hydrolyser ATP,
de se déplacer vers l’extrémité (+) des microfilaments (sauf la myosine VI qui se déplace à contre-sens).
La myosine interagit, non pas avec une actine quelconque, mais dans un environnement particulier avec des complexes à base d'actine à la bioarchitecture très sophistiqué.
Les nombreux facteurs qui sont nécessaires à la formation des réseaux à base d'actine travaillent de concert : les actines subissent une ségrégation spatiale et temporelle, contribuant ainsi à la création de compartiments intracellulaires avec une conception complexe et dynamique strictement régulée (Actin Nucleation and Elongation Factors: Mechanisms and Interplay 2009).
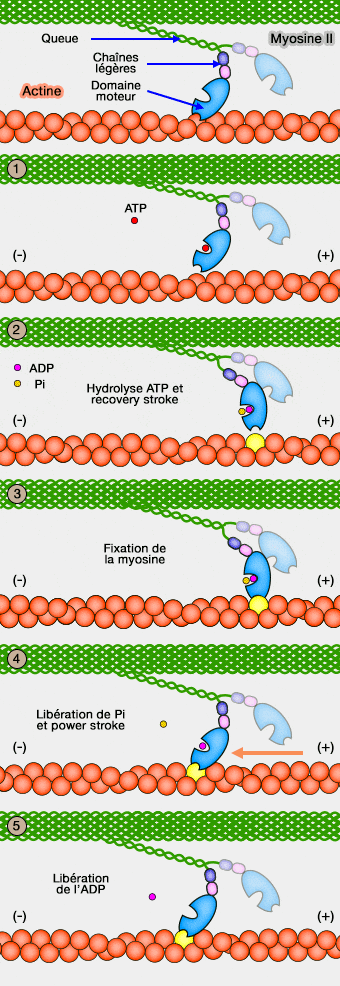
Liaison de la myosine II à l'actine
Vue d'ensemble
L'affinité de la myosine pour l'actine et l'ATP dépend du nucléotide (ATP ou ADP) et est antagoniste. Le cycle cross-bridge définit ces interactions cycliques entre actine et myosine.
Liaison simplifiée myosine-actine
(Figure : vetopsy.fr)
En l'absence d'ATP, la myosine est fixée fortement à l'actine : ce processus est de courte durée (configuration rigor), sauf lors de la mort (à l'origine de la rigidité cadavérique ou rigor mortis).
Lors de la contraction musculaire, cette liaison avec l'actine diminue l'affinité pour l'ATP 10 000 fois et vice-versa. La myosine ne reste liée à l'actine que pendant 10% du cycle ATPasique (10% de temps de travail)
Nous partons du stade où la myosine est fixée à l'actine.
U50 a fermé la " pince " qui enserre l'actine dans la fente entre U50 et L50. La poche nucléotidique est ouverte avec les deux commutateurs (SW1 et SW2) en position " ouverte "
1. Lors de la liaison de l'ATP avec la tête de la myosine, l'affinité à l'actine diminue fortement et provoque la dissociation de leur liaison.
L'ATP se lie dans la poche nucléotidique et U50 se déplace ce qui provoque l'ouverture des pinces de la tenaille qui libère l'actine et ferme SW1 sur l'ATP.
2. La tête de la myosine hydrolyse l'ATP en ADP et Pi qui restent liés et provoque un changement de conformation qui consiste en une rotation de la tête par rapport au cou (position " armée ", comme celle d'un pistolet : recovery stroke qui bloque le moteur).
Cette rotation du convertisseur (65°) et du domaine de liaison ferme SW2 sur l'ATP. Ce processus engage deux liaisons hydrogène qui déplace successivement l'hélice de relais (20°) et l'hélice SH1 (45°) liées au convertisseur. Le domaine du cou (lever arm) se déplace d'environ 60°. Le complexe myosine-ADP-Pi est stable quand les deux commutateurs sont fermés.
Ce modèle explique que la longueur du déplacement de la myosine (longueur de son pas) est proportionnelle à la longueur de son cou : plus long est le cou de la myosine, plus la vitesse de déplacement est rapide.
La myosine II à cou court se déplace de 8nm en 3 µm/s (force de 3-5 piconewtons), la myosine I de 10-14 nm et la myosine V de 36 nm.
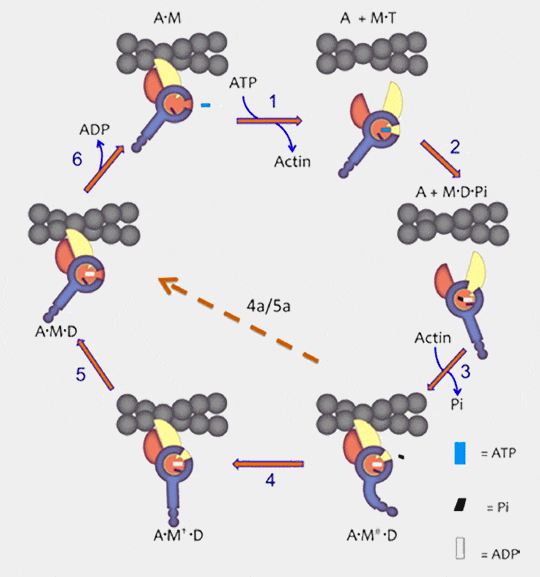
Cycle mécanochimique de la liaison myosine-actine
(Figure d'après Bloemink et Geeves)
3. La myosine se lie à l'actine après l'hydrolyse de l'ATP et provoque la torsion du convertisseur.
L'hydrolyse de l'ATP referme la tenaille U50-L50 (c'est-à-dire la fente) sur le filament d'actine par la rotation de U50 : U50 et L50 sont tous deux liés à l'actine. Cette rotation de U50 :
ouvre les commutateurs pour la libération de Pi,
provoque la torsion du convertisseur qui générera la force " élastique "
Normalement, la libération de Pi est très lente (phase la plus lente du cycle de l'ATP - demi-vie de l'ordre de 10 secondes pour le complexe myosine-ADP-Pi). En présence d'actine, cette libération est très rapide.
4. La torsion du convertisseur recourbe la tête d'actine qui déplace l'actine de 5-10 nm vers l'extrémité (+)(power stroke).
Remarque : 4a-5a (strain holding cross-bridge) : si la force est trop faible, le glissement n'a pas lieu. Le mouvement du segment bleu est de 5 à 100 fois moins grand selon les différentes myosines et dépend du degré de rotation, de la longueur du cou et de la rigidité du " ressort " de la structure.
5. Lorsque le convertisseur reprend sa position initiale, i;la tête de la myosine pivote et fait glisser l’actine, générant la contraction.une deuxième rotation de U50 ouvre la poche nucléotidique.
6. L'ADP peut s'échapper pour une nouvelle fixation de l'ATP et le cycle peut recommencer.




 power stroke).
power stroke).