


Système endo-lysosomal
Trafic des endosomes/lysosomes
Vue d'ensemble
- Vue d'ensemble du système endomembranaire
- Endosomes
- Lysosomes
- Trafic des endosomes/lysosomes
- Fusion et fission membranaire
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

La distribution spatio-temporelle des organites endo-lysosomaux dépend du transport généré essentiellement par les moteurs de microtubules (kinésines et dynéine), et par les moteurs du cytosquelette d'actine (myosines).
Vue d'ensemble du trafic
Le positionnement et le trafic des endosomes/lysosomes sont liés :
- aux protéines motrices des microtubules essentiellement, i.e. kinésines et dynéines,
- aux protéines motrices des filaments d'actine, i.e. les myosines,
- aux sites de contact membranaire (MCS).
Protéines motrices
Les protéines motrices diffèrent selon les éléments du cytosquelette.
Microtubules
1. Les protéines motrices des microtubules transportent les organites rapidement sur de grandes distances et de manière bidirectionnelle.
-
La plupart des kinésines déplacent les organites de l'extrémité moins vers l'extrémité plus (
 transport antérograde ou centrifuge).
transport antérograde ou centrifuge). - La dynéine provoque le déplacement en sens inverse, de l'extrémité plus vers l'extrémité moins (transport rétrograde ou centripète).
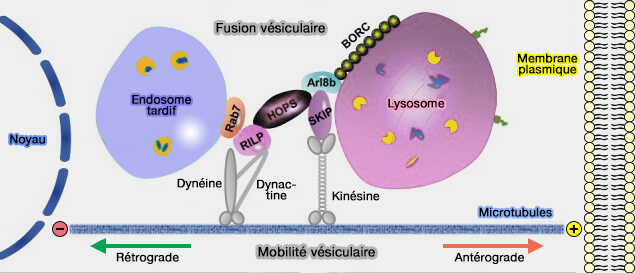
(Figure : vetopsy.fr d'après Khattern et coll)

L'équilibre entre le transport moteur de la dynéine rétrograde (centripète) et de la kinésine antérograde (centrifuge) détermine la direction nette du mouvement et la maturation des endosomes (Tug-of-war between dissimilar teams of microtubule motors regulates transport and fission of endosomes 2009).
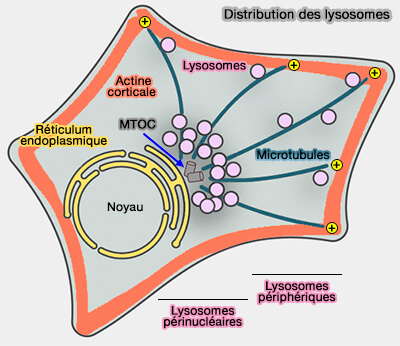
(Figure : vetopsy.fr d'après Cabukusta et coll)
2. Les réseaux de microtubules couvrent tout le cytosol.
Dans les cellules non polarisées, les microtubules sont distribués radialement (BORC/kinesin-1 ensemble drives polarized transport of lysosomes into the axon 2017 et Nucleation and Dynamics of Golgi-derived Microtubules 2015).
- Leurs extrémités négatives sont localisées au niveau du centre périnucléaire d'organisation des microtubules (MTOC).
- Leurs extrémités positives pointent vers la périphérie.
Dans les cellules polarisées, i.e. cellules épithéliales ou neurones, l'organisation est plus complexe.
- Des microtubules peuvent pointer leurs extrémités positives (+) vers le noyau.
- Le transport centrifuge ou centripète (-) est dépendant de microtubules spécifiques auxquels les organites sont attachés.
Filaments d'actine
1. Les moteurs des filaments d'actine, i.e. les myosines, interviennent sur de courtes distances et de manière lente (transport par le cytosquelette d'actine).
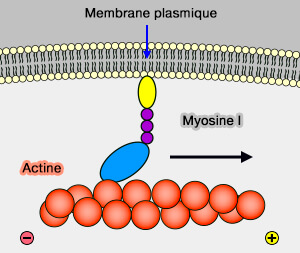
(Figure : vetopsy.fr)
2. Les fibres d'actine sont en majorité (mais pas exclusivement) concentrées sous la membrane plasmique pour déterminer la forme et la motilité des cellules (actine et cortex cellulaire).
Protéines adaptatrices des microtubules
1. Les protéines motrices, kinésines, dynéines et myosines ne se lient généralement pas directement aux organites sélectionnés, mais elles utilisent la plupart du temps :
- de petites GTPases qui, lorsqu'elles sont dans leur état actif, i.e. liées au GTP, marquent des organites définis et ensuite fonctionnent comme des échafaudages pour le recrutement des molécules impliquées dans les mécanismes de transport et de fusion sur des membranes cibles,
- des protéines effectrices des petites GTPases,
- des phosphoinositides ( positionnement des endosomes lié aux phosphoinositides).
2. Ce tableau est tiré de : Moving and Positioning the Endolysosomal System (2017).
| Organites | Moteurs | Adaptateurs et régulateurs |
|---|---|---|
| Endosomes précoces |
KIF5B | |
| KIF16B | ||
| Dynéine/Dynactine | ||
| Endosomes de recyclage |
KIF13A | |
| KIF13B | ||
| Dynéine | ||
| Endosomes tardifs/lysosomes |
KIF1A | |
| KIF1Bβ | ||
| KIF2A | ||
| KIF3A | KAP3 | |
| KIF5B | ||
| Dynéine/Dynactine | ||
| Autolysosomes (ALR) |
KIF5B | |
| Endosomes de recyclage (apicaux) |
KIF3A | |
| KIF3B | ||
| Endosomes transcytosiques |
KIF16B | Rab11 |
| Endosomes de signalisation (SARA) |
Dmel Klp98 (KIF16B) | |
| KIF5B | ||
| Dynéine/Dynactine | ||
| Mélanosomes | KIF5B | |
| Dynéine/Dynactine | ||
| Granules lytiques |
KIF5B | |
| Dynéine/Dynactine |
Sites de contact membranaire (MCS) des endosomes/lysosomes
Divers mécanismes de régulation garantissent que les interactions organites-moteur se produisent au bon endroit et au bon moment.
Un de ces mécanismes implique des contacts avec d'autres organites qui influencent les mouvements des organites endo-lysosomaux.
| Contacts organites /endosomes |
Contacts molécules / régulateurs |
|---|---|
| Réticulum endoplasmique (RE) et endosomes tardifs /lysosomes |
|
| Réticulum endoplasmique (RE) et tubules (rétromère) |
|
| Réticulum endoplasmique (RE) et endo-lysosomes |
|
| Appareil de Golgi et endosomes tardifs /lysosomes |
|
| Lysosomes et peroxysomes |
Le contrôle inter-compartimental du trafic endo-lysosomal est très complexe et implique :
MCS du réticulum
endoplasmique (RE)
1. Le réticulum endoplasmique (RE), omniprésent dans tout le cytoplasme, joue un rôle majeur dans le contrôle du mouvement et du positionnement global des autres organites.
2. Le réticulum endoplasmique contrôle, pour ce qui est des organites endo-lysosomaux, à travers un système complexe de protéines de liaison :
- leur positionnement global, par exemple, leur concentration dans la zone périnucléaire,
- leur motilité,
- leur fission,
- leur fusion.
Les MCS RE/MVB contribuent aussi à l'échange de lipides et de calcium.
Les sites de contact membranaire (MCS) sont traités dans des chapitres spéciaux, dont les MCS entre réticulum endoplasmique et endosomes ou corps multivésiculaires (MVE/MVB).
2. Ce système fonctionne dans les deux sens, car les endosomes influencent les mouvements du réticulum endoplasmique ( dynamique du RE liée aux endosomes).
MCS mitochondries/lysosomes
1. Les mitochondries interagissent aussi avec les lysosomes, et les lysosomes en mouvement peuvent extraire de fins tubules des mitochondries.
Des tubules minces similaires se sont avérés s'étendre des mitochondries aux points de contact avec le réticulum endoplasmique, via l'action de la petite GTPase Miro (Mitochondrial Rho GTPase 1), considérée maintenant comme une sous-classe des petites GTPases Ras (Understanding Miro GTPases: Implications in the Treatment of Neurodegenerative Disorders 2018), adaptateur pour le transport dépendant de KIF5B.
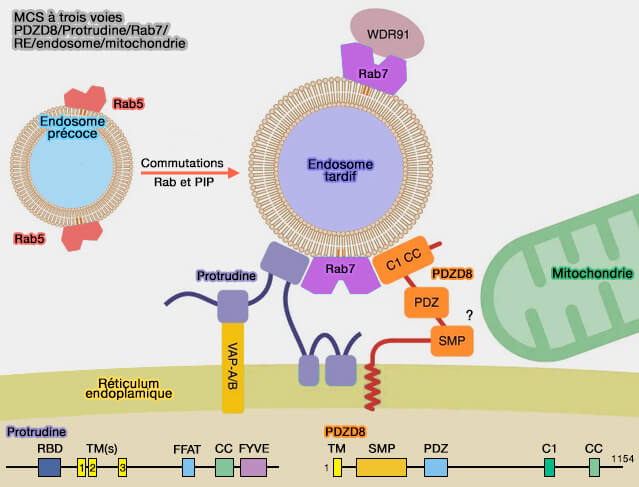
(Figure : vetopsy.fr d'après Elbaz-Alon et coll)
2. Est-ce que les sites de contact membranaire (MCS) à trois voies induites par PDZD8, i.e. protéine qui possède un domaine de transfert lipidique et interagit avec la protrudine, entre le réticulum endoplasmique (RE), les endosomes tardifs et les mitochondries pourraient être impliqués dans les deux processus ?
(Figure : vetopsy.fr d'après Starling et coll)
Toutefois, cette interaction complexe a essentiellement immobilisé les organites (PDZD8 interacts with Protrudin and Rab7 at ER-late endosome membrane contact sites associated with mitochondria 2020).
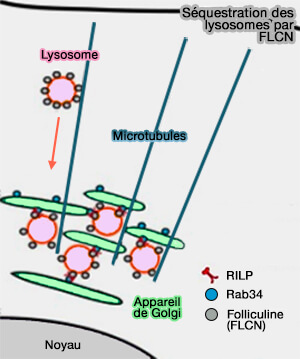
MCS appareil de Golgi/lysosomes
1. Les GTPases Rab34 et Rab36, principalement localisées sur l'appareil de Golgi, peuvent avoir un impact sur la distribution des lysosomes en favorisant leur regroupement périnucléaire (Rab36 regulates the spatial distribution of late endosomes and lysosomes through a similar mechanism to Rab34 2010).
2. La folliculine (FLCN), à ne pas confondre avec les hormones oestrogéniques, est impliquée dans la croissance et la prolifération cellulaire, la résistance au stress et l'autophagie.
FLCN contrôle la localisation périnucléaire des lysosomes lors de privation.
- Elle se lie directement, via son domaine DENN C-terminal, avec l'effecteur RILP de Rab34 (Folliculin directs the formation of a Rab34–RILP complex to control the nutrient‐dependent dynamic distribution of lysosomes 2016).
- La folliculine est associée au syndrome de Birt-Hoge-Dubé (BHD), syndrome caractérisé par des lésions cutanées, des tumeurs rénales et des kystes pulmonaires qui peuvent conduire à un pneumothorax.
Trafic des endosomes/lysosomes : trafic antérograde (centrifuge)
Biologie cellulaire et moléculaireMembrane plasmiqueNoyauCytoplasmeMitochondriesSystème endomembranaireRéticulum endoplasmiqueAppareil de GolgiEndosomes/LysosomesPeroxysomesProtéasomesCytosquelettemicrofilaments d'actineFilaments intermédiairesMicrotubulesMatrice extracellulaireReproduction cellulaireBiochimieTransport membranaireMoteurs moléculairesVoies de signalisation