


Lipides
Gouttelettes lipidiques
(Lipid droplets ou LD)
Sites de contact membranaires (MCS)
- Biochimie
- Chimie organique
- Bioénergétique
- Composition de la matière vivante
- Composés organiques
- Protides
- Acides nucléiques
- Glucides
- Lipides
- Coenzymes
- Hormones
- Composés inorganiques
- Composés organiques
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

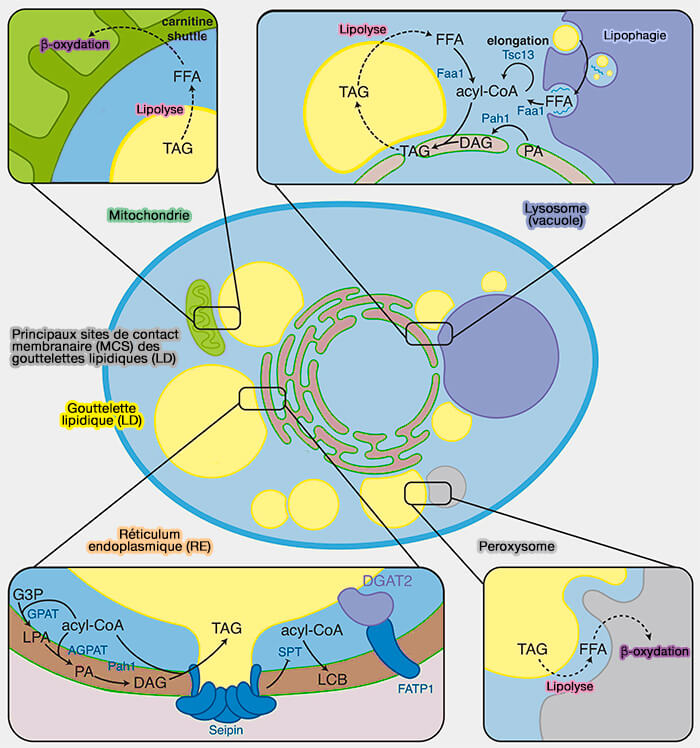
Les gouttelettes lipidiques sont en contact avec différents organites dans des réseaux cellulaires dynamiques et coopératifs qui maintiennent l'homéostasie lipidique
Les sites de contact membranaire (MCS) sont des régions d'apposition étroite (≤ 30 nm) entre deux organites, établissant des microdomaines qui permettent un échange de matériel.

Les progrès récents de la biologie des MCS ont révélé leurs rôles clés dans les échanges inter-organites. Les recherches sont très actives, i.e. les connaissances évoluent rapidement et doivent être mises à jour continuellement.
Vue d'ensemble des MCS des LD
Les MCS des LD sont des sites de transfert de constituants cellulaires vers ou depuis les LD pour (Lipid Droplet-Organelle Contact Sites as Hubs for Fatty Acid Metabolism, Trafficking, and Metabolic Channeling 2021) :
- la mobilisation d'énergie en réponse aux limitations en nutriments,
- leur nucléation et leur expansion,
- leur autophagie.
(Figure : vetopsy.fr d'après Freyre et col)
1. Les gouttelettes lipidiques (LD) sont en contact avec différents organites impliqués dans le métabolisme lipidique (Applying systems-level spectral imaging and analysis to reveal the organelle interactome 2017) :
- le réticulum endoplasmique (RE),
- les mitochondries,
- les peroxisomes,
- les endosomes/lysosomes,
- l'appareil de Golgi,
- le noyau,
- les autres LD.
Remarque : de nombreux MCS sont tripartites, i.e. entre plusieurs organites, comme par exemple les MCS mitochondries-RE-LD pour le stockage des triglycérides (TG).
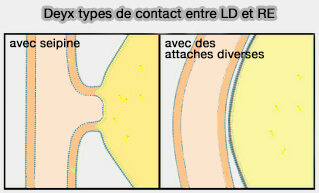
2. Les MCS LD-organites présentent des caractéristiques distinctives qui les distinguent de tous les autres sites de contact membranaire (MCS).
a. À cause des propriétés physiques et de la composition des LD, les protéines de transport conventionnelles intégrées dans les bicouches lipidiques qui participent au transfert des lipides et empêchent leur fusion, n'entrent pas en ligne de compte (A different kind of love – lipid droplet contact sitesSchuldiner et Bohnert, 2017).
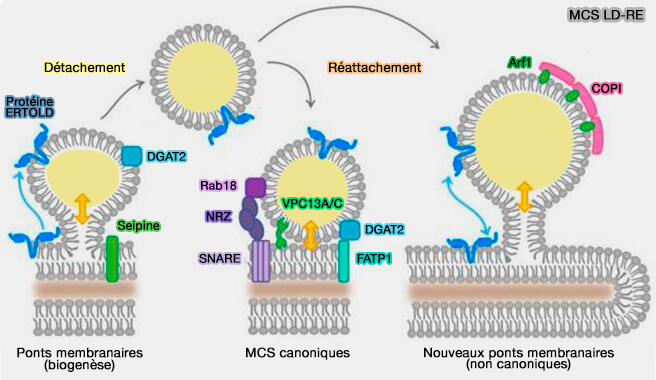
- Ce n'est pas le cas des MCS avec LD, en particulier au stade de la biogenèse, où les deux membranes (double couche pour l'organite et hémi-couche pour les LD) fusionnent effectivement par des intermédiaires d'hémifusion.
- Des ponts membranaires transitoires entre des gouttelettes lipidiques et d'autres organites, suggèrent des propriétés uniques pour ces MCS.
b. Au lieu de cela, des protéines spécialisées, telles que les lipases, les LTP (Lipid Tranfer Protein), mais aussi les MCS eux-mêmes permettent ce transfert (Dynamics and functions of lipid droplets 2019).
(Figure : vetopsy.fr d'après Renne et Hariri)
3. Les complexes protéiques d'attache, qui jouent un rôle clé dans l'établissement et le maintien des sites de contact des organites, sont mal connus quant à leur identité, leurs mécanismes de régulation et leurs fonctions restent pour la plupart inconnus.
4. Le cytosquelette joue aussi un rôle essentiel dans les MCS ( LD et cytosquelette).
LD et cytosquelette).
MCS LD-RE
Les MCS RE/LD sont uniques en leur genre, affichant souvent une continuité membranaire entre les deux organites, ce qui les différencient des autres MCS, de part la biogenèse des LD qui est développée dans les chapitres correspondants ( biogenèse des LD).
Les LD restent le plus souvent en contact avec le RE ( détachement ou non des LD).
(Figure : vetopsy.fr d'après Salo et coll)
1. Ce contact serait du en partie à l'interaction avec la seipine (A Unique Junctional Interface at Contact Sites Between the Endoplasmic Reticulum and Lipid Droplets 2021).
Les protéines de transfert de lipides comme VPS13 ou ORP5/8 pourraient également, en plus de leur rôle dans l'expansion des LD, servir d'attache.
2. Cependant, bien d'autres protéines d'attache pourraient intervenir à ce niveau (Moving out but keeping in touch: contacts between endoplasmic reticulum and lipid droplets 2019 et Tethering Fat: Tethers in Lipid Droplet Contact Sites 2020 pour une liste).
a. MOSPD2, une protéine de la famille VAP (VAMP-Associated Protein), se distingue des autres membres de la famille VAP par la présence d'un domaine CRAL-TRIO qui se lie aux LD par une hélice hydrophobe (MOSPD2 is an endoplasmic reticulum–lipid droplet tether functioning in LD homeostasis 2022).
(Figure : vetopsy.fr d'après Vieyres et Pietschmann)
b. DGAT2 pourrait s'associer à FATP1 (Fatty Acid Transport Protein 1), une acyl-CoA synthétase localisée aussi dans la membrane plasmique (The FATP1–DGAT2 complex facilitates lipid droplet expansion at the ER–lipid droplet interface 2012 et The role of FATP1 in lipid accumulation: a review 2021).
c. Rab18 est recruté à la surface des LD par le complexe métazoaire NAG-RINT1-ZW10 (NRZ) qui correspond à la protéine d'amarrage (tethering protein) Dsl1 de la levure, associé au RE et aux complexe SNARE, i.e. syntaxine18, Use1 et Sec20/BNIP1 pour semble-t-il une régulation de la biogenèse (Rab18 promotes lipid droplet (LD) growth by tethering the ER to LDs through SNARE and NRZ interactions 2018).
Ce complexe est efficace dans les adipocytes, mais beaucoup moins dans d'autres cellules, ce qui pourrait laisser supposer que certaines attaches sont spécifiques au contexte.
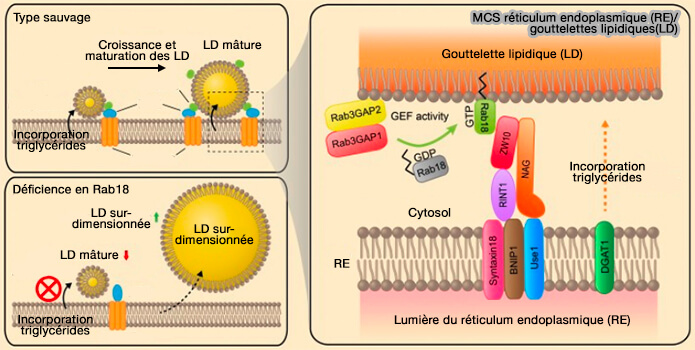
(Figure : vetopsy.fr d'après Xu et coll)
Remarque : Snx14/Mdm1, une nexine résidant dans le RE associée à l'ataxie cérébelleuse SCA20, se localise dans les contacts RE-LD indépendamment de la seipine, où elle favoriserait la croissance et la maturation du LD (Cerebellar ataxia disease–associated Snx14 promotes lipid droplet growth at ER–droplet contacts 2019).
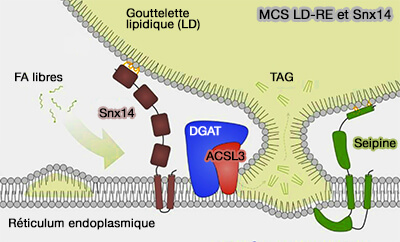
(Figure : vetopsy.fr d'après Liu et col)
Elle interviendrait aussi dans la lipophagie dans les MCS LD-lysosomes (Mdm1 maintains endoplasmic reticulum homeostasis by spatially regulating lipid droplet biogenesis 2019).
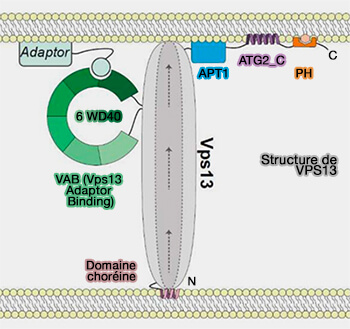
3. VPS13A et VPS13C se sont récemment révélés être des protéines de transfert de glycérolipides à haute capacité qui se localisent dans les LD (The Vps13 Family of Lipid Transporters and Its Role at Membrane Contact Sites (2021).
VPS13 est étudié dans le chapitre des LTP ou protéines de transfert des lipides.
a. VPS13A et VPS13C se localisent aux contacts RE-LD via leurs motifs FFAT qui lient les VAP (VAMP-Associated Protein) au niveau du RE et des hélices amphipatiques (AH) qui ciblent les LD (VPS13A and VPS13C are lipid transport proteins differentially localized at ER contact sites 2018).
- VPS13A améliore la localisation des LD au RE dans les cellules traitées avec de l'acide oléique, un inducteur de la formation de LD, i.e. en absence de VPS13A le nombre de gouttelettes lipidiques est augmenté
- VPS13A se localise également sur les gouttelettes lipidiques et affecte la motilité des gouttelettes lipidiques.
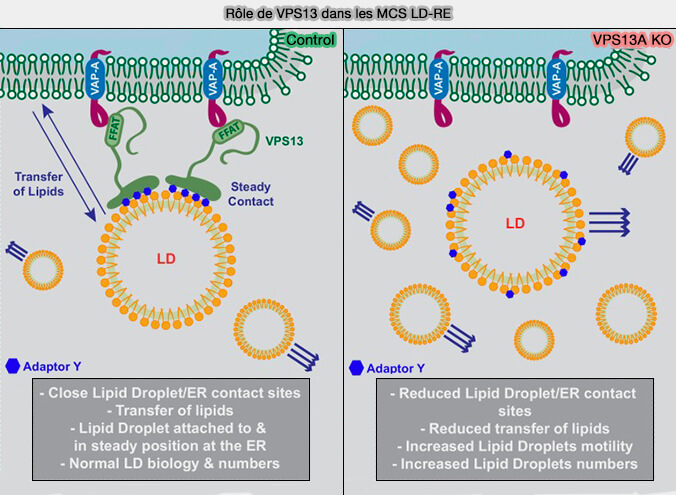
(Figure : vetopsy.fr d'après Yeshaw et coll)
Remarque : VPS13A, mais aussi VPS13D avec le complexe ESCRT, intergit aussi avec les mitochondries via son domaine C-terminal.
Dans les cellules appauvries en VPS13A, le nombre de MCS RE-mitochondries est diminué, les mitochondries sont fragmentées et la mitophagie est abaissée.
b. VPS13A est nécessaire pour établir des sites de contact membranaire entre divers organites afin de permettre le transfert de lipides requis pour les processus liés aux mitochondries et aux gouttelettes lipidiques. (Human VPS13A is associated with multiple organelles and influences mitochondrial morphology and lipid droplet motility 2019).
MCS LD-lysosomes
La formation de sites de contact membranaire (MCS) entre les LD et les lysosomes, i.e. vacuoles chez la levure correspondant aux vCLIP (vaCuole LIPid droplet contact sites), joue un rôle direct et indirect dans l'autophagie des LD (lipophagie).
Les trois formes de lipophagie sont étudiées dans un chapitre spécifique.
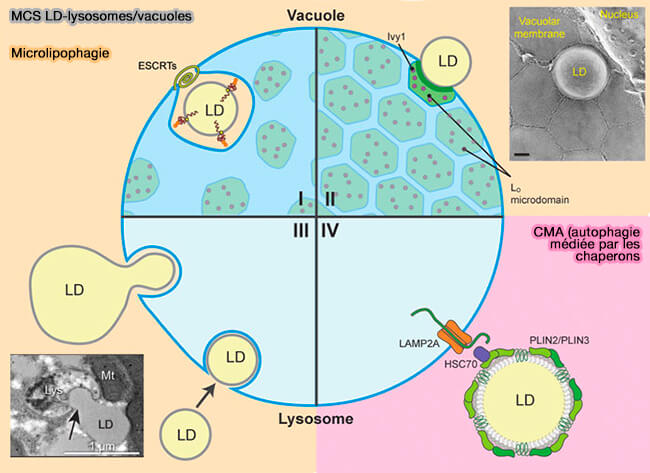
(Figure : vetopsy.fr d'après Liao et et coll)
Les MCS LD-lysosomes sont essentiels dans :
- la microautophagie ou microlipophagie LD (µLP) par une absorption partielle (fragmentaire) ou totale de la LD,
- par la CMA, l'autophagie médiée par les chaperons (Chaperone-Mediated Autophagy).
MCS LD-Golgi
Le réseau trans-Golgi (TGN) est le site majeur de tri des protéines et des lipides vers différents sites cellulaires (Hello From The Other Side: Membrane Contact Of Lipid Droplets With Other1 Organelles and Subsequent Functional Implications 2020).
(Figure : vetopsy.fr d'après Dziurdzik et coll)
1. Toutefois, les MCS LD-Golgi ne sont pas bien connues, même si les LD et le Golgi peuvent se rapprocher (Applying systems-level spectral imaging and analysis to reveal the organelle interactome 2017) .
Il semblerait que VPS13B pourrait servir d'attache dans ces MCS chez l'homme via RAB6 pour transférer les lipides afin de maintenir l'intégrité du Golgi lors de stéatose (Trans-Golgi network-lipid droplet contacts maintain the TGN integrity and function via lipid transfer activities of VPS13B 2020).
2. D'autres protéines pourraient intervenir :
- ORP5/8, comme dans les MCS LD-RE,
- les golgines peut-être (The Golgin Family of Coiled-Coil Tethering Proteins 2016).
MCS LD-noyau
Les MCS LD-noyau sont mal connues ( biogenèse des nLD).
MCS LD-LD
Les MCS LD-LD font intervenir les protéines CIDE, Plin1 et Rab8 pour la fusion des LD ( expansion et fusion des LD).
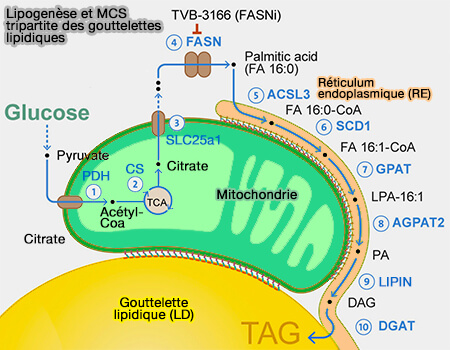
MCS LD-mitochondries et LD-peroxysomes
BiochimieChimie organiqueBioénergétiqueProtidesGlucidesLipidesAcides grasLipides vraisLipides vrais simplesLipides vrais complexesLipoïdesCholestérolGlycéridesPhospholipidesTrafic non vésiculaire et LTPDigestion et absorption entérocytaire des lipidesLipoprotéinesApolipoprotéinesGouttelettes lipidiques (LD)EnzymesCoenzymesVitaminesHormonesComposés inorganiquesTransport membranaireMoteurs moléculairesVoies de signalisation