


Système endo-lysosomal
Complexe ESCRT
Vue d'ensemble
- Vue d'ensemble du système endomembranaire
- Endosomes
- Vue d'ensemble des endosomes
- Endosomes précoces
- Endosomes de recyclage
- Vésicules intraluminales (ILV) et endosomes ou corps mutivésiculaires (MVE/MVB)
- Endosomes tardifs
- Système endo-lysosomal et phosphoinositides
- MCS (sites de contact membranaire) RE/endosomes/lysosomes
- Lysosomes
- Trafic des endosomes/lysosomes
- Fusion et fission membranaire
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

Le complexe ESCRT (Endosomal Sorting Complex Required for Transport ou complexe de tri endosomal requis pour le transport) est composé de 5 complexes impliqués pendant les évènements de remodelage de la membrane cellulaire.
Le complexe ESCRT comprend :
- ESCRT-0,
- ESCRT-I,
- ESCRT-II,
- ESCRT-III,
- Vps4 (Vacuolar Protein Sorting 4) qui contient un domaine catalytique AAA+ ATPase.
Les protéines constituant la machinerie ESCRT font partie d’une classe plus large de protéines appelées protéines VPS ou vacuolar protein sorting (Morphological Classification of the Yeast Vacuolar Protein Sorting Mutants: Evidence for a Prevacuolar Compartment in Class E vps Mutants 1992).
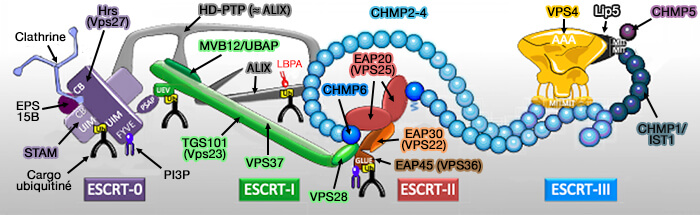
(Figure : vetopsy.fr d'après Christ et Hill)
Rôles du complexe ESCRT
Le complexe ESCRT a plusieurs rôles qui impliquent des processus de remodelage cellulaire, et en particulier des déformations membranaires (Reverse-topology membrane scission by the ESCRT complexes 2017 et Dynamic and elastic shape transitions in curved ESCRT-III filaments 2017).
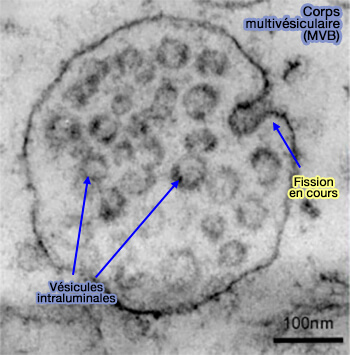
(Figure : vetopsy.fr d'après Chivet et coll)
1. Le complexe ESCRT forme les vésicules intraluminales (ILV) des endosomes ou corps multivésiculaires - MVE/MVB - (ESCRT-dependent cargo sorting at multivesicular endosomes 2017).
Le complexe ESCRT est impliqué dans la formation de la majorité des vésicules extracellulaires (EV) :
- les exosomes, i.e. vésicules dérivant des MVE/MVB qui relarguent leur contenu dans le milieu extracellulaire,
- les ectosomes, i.e. vésicules formées à partir de la membrane plasmique qui se détachent dans le milieu extracellulaire.
2. Le complexe ESCRT est engagé dans :
- l'abcission de la membrane plasmique lors de la cytokinèse, dernière étape de la division cellulaire (Knowing when to cut and run: mechanisms that control cytokinetic abscission 2013 et Membrane Fission Reactions of the Mammalian ESCRT Pathway 2013 et Cytokinetic Abscission: Molecular Mechanisms and Temporal Control 2014).
- la fission et la reformation de l'enveloppe nucléaire (ESCRT-III controls nuclear envelope reformation 2015).
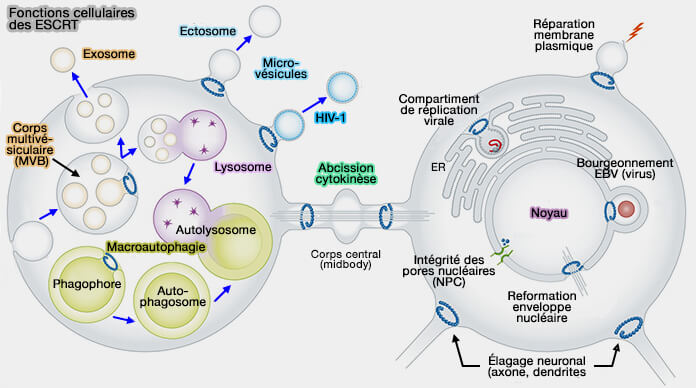
(Figure : vetopsy.fr d'après Hurley)
3. Le complexe ESCRT joue un rôle dans la réparation des membranes :
- plasmique (ESCRT Machinery Is Required for Plasma Membrane Repair 2014),
- lysosomale et nucléaire (ESCRT III repairs nuclear envelope ruptures during cell migration to limit DNA damage and cell death 2016),
- la fermeture des autophagosomes (ESCRTs and Fab1 Regulate Distinct Steps of Autophagy 2007).
4. Le complexe ESCRT intervient dans :
- l'élagage neural (pruning en anglais), i.e. clivage et élimination des neurites inutiles (An ESCRT module is required for neuron pruning 2015)
- le remodelage des épines dendritiques (Rôle de CHMP2B et du complexe ESCRT-III dans le remodelage dans membranes cellulaires: cas des épines dendritiques 2017).
6. Enfin, le complexe ESCRT intervient dans le bourgeonnement de certains virus enveloppés (VIH par exemple) pour catalyser la fission membranaire.

Les sous-unités du complexe ESCRT se lient à PI(3)P (The class III phosphatidylinositol 3-kinase Vps34 in Saccharomyces cerevisiae 2016 et Rab GTPases in organelle biogenesis and transport).
Structure générale des ESCRT
On trouve plus d'une vingtaine d'ESCRT (Cellular Functions and Molecular Mechanisms of the ESCRT Membrane-Scission Machinery 2017).
Le coeur du complexe ESCRT comprend plusieurs sous-complexes et plusieurs protéines.
1. Les ESCRT sont composés de trois sous-unités biochimiquement distinctes.
a. ESCRT-I et ESCRT-II contribuent principalement :
- au tri des protéines,
- au recrutement de ESCRT-III par l'intermédiaire de l'association avec des facteurs de ciblage en amont. Ce recrutement dépend essentiellement des protéines de domaine ESCRT-I et des protéines à domaine BRO1 comme ALIX et HD-PTP.
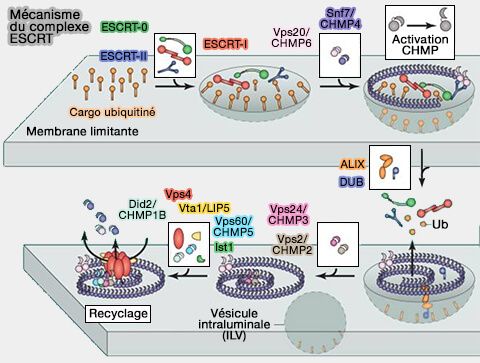
(Figure : vetopsy.fr d'après Wollert et coll)
b. ESCRT-IIII est considéré comme le principal moteur du remodelage et de la scission membranaire.
- il existe sous forme de monomères solubles en solution.
- Lorsqu'il est activé par ESCRT-II ou ALIX, ESCRT-III se polymérise sous forme de filaments spiralés de morphologie, de composition et de stoechiométrie variables, qui déforment la membrane vers l’intérieur de la lumière de l’endosome, et permet ainsi la formation des vésicules intraluminales (ILV) et des endosomes ou corps multivésiculaires (MVE/MVB).
Le désassemblage de l'ESCRT-III nécessite l'action du dernier composant central de l'ESCRT, l'hexamère AAA+ ATPase VPS4.
2. En outre, ESCRT-0, par son domaine DUIM et ses deux motifs motifs UIM, est un complexe qui initie le transport des cargos ubiquitinés dans les ILV, i.e. l'ubiquitine est le principal signal de cette voie.
Toutefois, les ESCRT-I (domaine UEV) et ESCRT-II (domaine GLUE) ainsi qu'ALIX (QRE et ERE) contiennent aussi des motifs de liaison à l'ubiquitine qui participent au tri des cargos (How Ubiquitin Functions with ESCRTs 2011).
ESCRT-0 et cargos ubiquitinés et ESCRT-I
Biologie cellulaire et moléculaireMembrane plasmiqueNoyauCytoplasmeMitochondriesSystème endomembranaireRéticulum endoplasmiqueAppareil de GolgiEndosomesLysosomesPeroxysomesProtéasomesCytosquelettemicrofilaments d'actineFilaments intermédiairesMicrotubulesMatrice extracellulaireReproduction cellulaireBiochimieTransport membranaireMoteurs moléculairesVoies de signalisation