


Autophagie
Macroautophagie
Macroautophagie sélective
Modèles
- Vue d'ensemble du système endomembranaire
- Autophagie
- Système UPS (Système UPS (Ubiquitine-protéasome))
- Fusion et fission membranaire
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

Plusieurs modèles d'autophagie sélective peuvent être envisagés.
L'identification des récepteurs des cargos a initialement prédit un modèle linéaire simple d'autophagie sélective, dans laquelle le cargo est reconnu par des récepteurs d'autophagie spécifiques (SAR) qui recrutent davantage de phagophores contenant des LC3/Atg8 pour faciliter la séquestration du cargo.
Modèle chez la levure
La levure fait appel à Atg11.
Les différences architecturales entre Atg11 avec ses domaines coiled-coil (CC) parallèles et Atg17 avec ses CC antiparallèles qui forment des dimères en forme de S expliqueraient la variation de l'organisation du PAS pour l'autophagie sélective et non sélective.

Le modèle de la levure avec Atg11 est traité dans le chapitre correspondant.
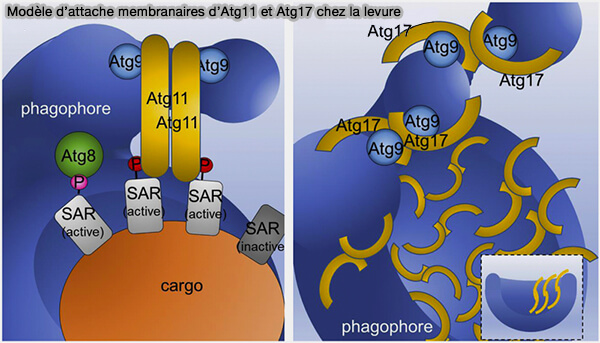
(Figure : vetopsy.fr d'après Zientara-Rytter et Subramani)
Modèle chez les mammifères
À l'heure actuelle, le modèle serait le suivant.
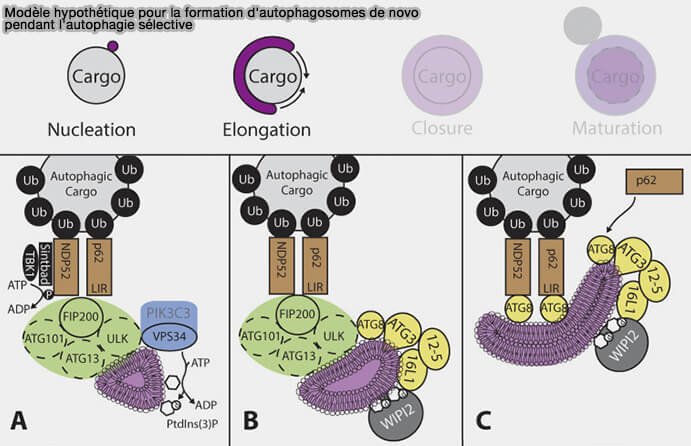
(Figure : vetopsy.fr d'après Melia et coll)
Les récepteurs de l'autophagie, par exemple p62 et NDP52, interagissent avec la sous-unité FIP200 du complexe ULK pour initier la formation d'autophagosomes de novo autour du cargo (Spatiotemporal Control of ULK1 Activation by NDP52 and TBK1 during Selective Autophagy 2019).
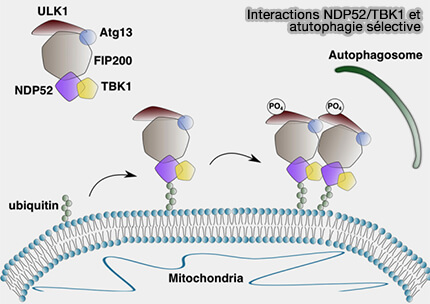
(Figure : vetopsy.fr d'après Alers et coll)
Le recrutement d'ULK permettra celui de VPS34 du complexe PI3KC3-CI responsable de la production de PI(3)P sur les omégasomes.
Exemple de NDP52
1. La phosphorylation médiée par TBK1 (Tank-Binding Kinase 1) de NDP52 (Nuclear dot protein 52) stimule l'interaction NDP52-FIP200 (Spatiotemporal Control of ULK1 Activation by NDP52 and TBK1 during Selective Autophagy 2019).
- NDP52/TBK1 cible le complexe ULK au cargo pour initier l'autophagie en l'absence de LC3.
- Dans les mitochondries, l'ubiquitination a été suivie par l'activation de TBK1, conduisant au recrutement de FIP200 et des récepteurs autophagiques comme l'optineurine, et plus tard, celui d'ATG13 (Selective Autophagy of Mitochondria on a Ubiquitin-Endoplasmic-Reticulum Platform 2019).
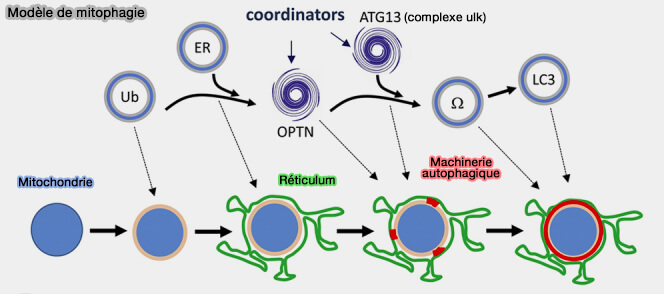
(Figure : vetopsy.fr d'après Zachari et coll)
Remarque : TBK1 joue aussi d'autres rôles (TBK1 (TANK-binding kinase 1)-mediated regulation of autophagy in health and disease 2021).
- La phosphorylation de STX17 sur S202 permet la translocation des sites de nucléation de l'appareil de Golgi aux phagophores (
 Rôle de STX17 dans l'initiation de l'autophagie).
Rôle de STX17 dans l'initiation de l'autophagie). - La phosphorylation par TBK1 de LC3C et GABARAPL2 régule leur délipidation en perturbant spécifiquement la capacité des protéines ATG4 ( délipidation des Atg8).
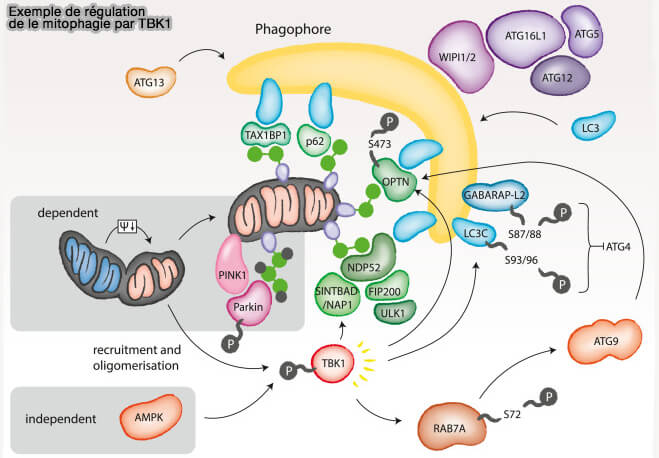
(Figure : vetopsy.fr d'après Kumar et coll)
Exemple de p62/SQTM1

(Figure : vetopsy.fr d'après Liu et coll)
1. La protéine p62/SQSTM1 (ubiquitin-binding protein 62/Sequestosome-1) est capable de reconnaître et de recruter spécifiquement un cargo à dégrader par le biais de sites d’ubiquitination (SQSTM1/p62: A Potential Target for Neurodegenerative Disease 2019).
- p62 dispose d’un domaine de reconnaissance des ubiquitines et d'un motif LIR de liaison à la LC3, le tout lui permettant d’adresser le contenu à l’autophagosome (SQSTM1/p62 and Hepatic Mallory-Denk Body Formation in Alcohol-Associated Liver Disease 2023).
- De plus, la p62 est également dégradée lors de la dernière étape de l’autophagie, i.e. son accumulation témoigne donc d’un arrêt du flux autophagique.
Remarque : p62 est aussi impliquée dans le système ubiquitine-protéasome ou UPS (p62 links the autophagy pathway and the ubiqutin–proteasome system upon ubiquitinated protein degradation 2016).
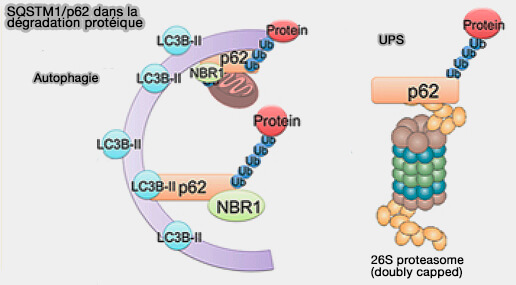
(Figure : vetopsy.fr d'après Ma et coll)
2. L'interaction p62-FIP200 nécessite le domaine LIR de p62 (FIP200 Claw Domain Binding to p62 Promotes Autophagosome Formation at Ubiquitin Condensates 2019).
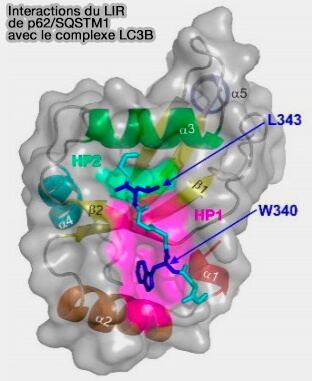
(Figure : vetopsy.fr d'après Wesch et coll)
a. Les résidus désordonnés 326–380 de p62 interagissent directement avec la région C-terminale (CTR) de FIP200.
- Le CTR FIP200 contient un domaine globulaire dimérique, appelé " claw " pour griffe ou serre en français à cause de sa forme.
- L’interaction de p62 avec FIP200 est médiée par une poche chargée positivement dans la griffe, renforcée par la phosphorylation de p62, mutuellement exclusive avec la liaison de p62 à LC3B, et elle favorise la dégradation de la cargaison ubiquitinée par autophagie.
b. De plus, le recrutement du CTR de FIP200 ralentit la séparation de phase des protéines ubiquitinées par p62.
3. L'interaction p62/FIP200 pourrait être complétée par celle de LC3B, indiquant un ordre séquentiel de partenaires de liaison p62 (FIP200 Claw Domain Binding to p62 Promotes Autophagosome Formation at Ubiquitin Condensates 2019).
- Ainsi, les protéines LC3/GABARAP peuvent être recrutées dans des mitochondries endommagées indépendamment de leur liaison aux récepteurs autophagiques.
- Elles stimulent alors les récepteurs autophagiques, i.e. OPTN et NDP52, d'une manière LIR-dépendante, mais ubiquitine-indépendante, ce qui facilite le recrutement supplémentaire des récepteurs de l'autophagie et l'expansion de la membrane autophagique autour du substrat sélectif (The mechanisms and roles of selective autophagy).
- Une boucle feedforward positive dépendante de LC3/GABARAP permettrait l'expansion du phagophore et la mitophagie.
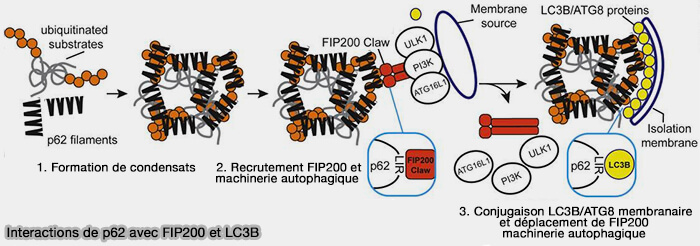
(Figure : vetopsy.fr d'après Turco et coll)
Remarque : NBR1 (neighbor of BRCA1 gene 1) est souvent associé à p62 dans l'agrégophagie.
- NBR1 est le principal récepteur de l’autophagie chez les plantes qui ne contiennent pas p62.
- Les SAR de la levure Atg19 et Atg34 sont des homologues de NBR1.
- La plupart des animaux contiennent à la fois NBR1 et p62.
NBR1 serait impliqué dans les maladies humaines telles que les protéinopathies et le cancer.

Vous pouvez lire : NBR1: The archetypal selective autophagy receptor (2022)) qui détaille sa structure et ses rôles.
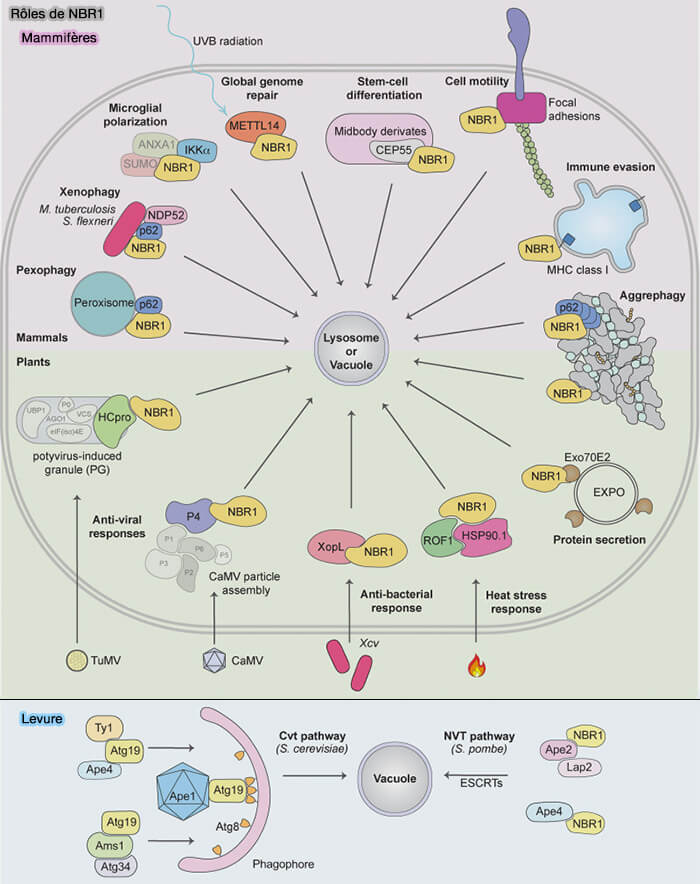
(Figure : vetopsy.fr d'après Rasmussen et coll)
Modèle général
La microscopie a démontré que l'initiation du phagophore semble se produire sur plusieurs sites de mitochondries étroitement liés au réticulum endoplasmique (Selective Autophagy of Mitochondria on a Ubiquitin-Endoplasmic-Reticulum Platform 2019).
Un modèle pour la biogenèse de l'autophagosome dans l'autophagie sélective pourrait être le suivant.
- L'ubiquitination des protéines cargo agit comme un signal " eat-me " pour initier l'activation de TBK1 et la liaison des récepteurs autophagiques, conduisant au recrutement de FIP200 et à celui de la machinerie autophagique pour faciliter la lipidation des protéines ATG8.
- ATG8 peut alors recruter d'autres récepteurs autophagiques.
- La force des interactions des récepteurs de cargos avec les protéines de la famille ATG8 localisées à la membrane facilite le transport du cargo en excluant probablement tout matériau non ciblé par la séquestration dans les autophagosomes.
Différences entre macroautophagie en vrac (" bulk autophagy ")
et macroautophagie sélective
Nucléation
1. Dans l’autophagie en vrac, l’activation d’ULK1 conduit à l’accumulation des complexes ULK1/2 et PI3KC3-CI sur les extensions membranaires du réticulum endoplasmique (RE) DFCP1-positives (omégasomes), ce qui crée une plate-forme pour la nucléation du phagophore.
- Une étape essentielle est la synthèse de PI(3)P sur le phagophore.
- Cela induit un recrutement d’autres protéines d’autophagie de base impliquées dans la nucléation du phagophore et la lipidation des protéines ATG8.
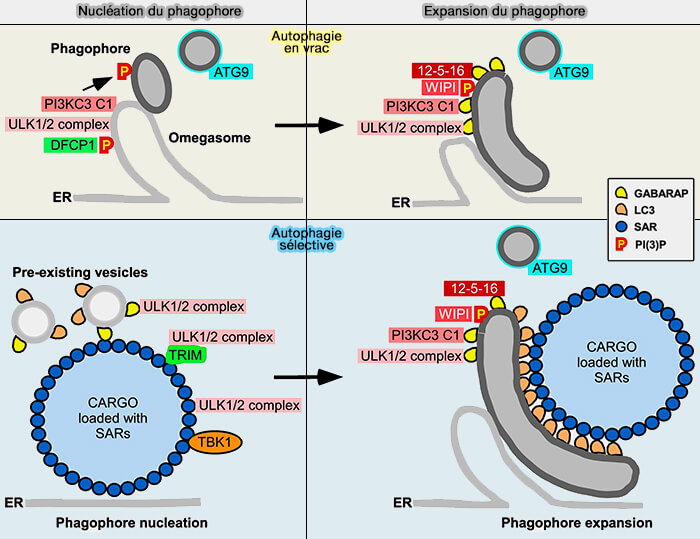
(Figure : vetopsy.fr d'après Johansen et Lamark)
2. Dans l’autophagie sélective, la nucléation du phagophore se produit à partir d’un mPAS formé in situ au niveau du cargo recouvert de récepteurs sélectifs de l'autophagie (SAR).
a. Comme dans l’autophagie en vrac, le processus de nucléation dépend d’un recrutement des complexes ULK1/2 et PI3KC3-CI et plusieurs processus d’autophagie sélective dépendent d’un recrutement de TBK1.
b. La façon dont les complexes initiaux sont recrutés peut différer entre les voies d’autophagie sélectives, mais cela peut se faire par :
- une liaison directe au récepteur de l'autophagie sélective (SAR),
- via d’autres protéines recrutées par la sélection du cargo, par exemple, les ligases E3 de la famille TRIM.
c. Potentiellement, les protéines d’autophagie de base avec un motif LIR peuvent également être recrutées via des vésicules préexistantes contenant du GABARAP lipidé.
Expansion du phagophore
Dans les deux types de macroautophagie, l’assemblage des protéines d’autophagie centrales sur le bord du phagophore en croissance est stabilisé par les interactions du motif LIR avec les GABARAP lipidés.
Dans l’autophagie sélective, la formation de multiples interactions LIR entre le SAR et les ATG8 lipidés à la surface interne du phagophore est essentielle pour l’amarrage de la cargaison au phagophore en croissance.
Retour à la macroautophagie
Biologie cellulaire et moléculaireConstituants de la celluleSystème endomembranaireRéticulum endoplasmiqueAppareil de GolgiEndosomesLysosomesPeroxysomesAutophagieMacroautophagieMicroautophagieAutophagie chaperonnes (CMA)Autophagie non canoniqueProtéines ATGSystème UPS (Ubiquitine-Protéasome)Transport membranaireTrafic vésiculaireFusion/fission membranaireEndocytoseVoie sécrétoireMoteurs moléculairesVoies de signalisation