


Système endo-lysosomal
Réticulum endoplasmique
Sites de contact membranaire (MCS) du RE
Endosomes multivésiculaires (MVE/MVB) et lysosomes
3. Dynamique du RE
- Vue d'ensemble du système endomembranaire
- Endosomes
- Lysosomes
- Trafic des endosomes/lysosomes
- Fusion et fission membranaire
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

Les sites de contact membranaire (MCS) du réticulum endoplasmique avec les endosomes/lysosomes sont utilisés pour contrôler la dynamique du RE par l'extension de microtubules (auto-stop ou hitchhiking en anglais).
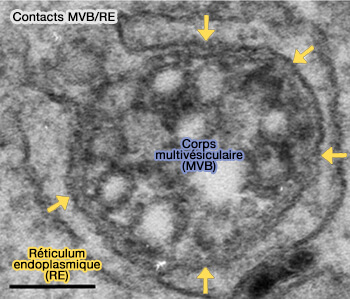
(Figure : vetopsy.fr d'après Wong et coll)
Les fonctions principales des MCS du réticulum endoplasmique sont essentielles, en particulier pour les endosomes ou corps multivésiculaires (MVE/MVB) et les lysosomes pour :
- l'échange de lipides par formation de gouttelettes lipidiques (Lipid Droplets ou LD),
- l'échange de calcium,
- la signalisation des facteurs de croissance (The EGFR odyssey - from activation to destruction in space and time 2017),
- le trafic des endosomes le long des microtubules,
- le positionnement des endosomes,
- la fission membranaire,
- la fusion membranaire,
- la dynamique du réticulum endoplasmique,
- la croissance des neurites,
- la formation d'autophagosomes,
- la signalisation du récepteur tyrosine kinase…
Dynamique du réticulum
endoplasmique

La dynamique du réticulum endoplasmique est étudiée dans un chapitre spécial.
1. L'extension des tubules du RE est provoquée par :
- les protéines motrices, i.e. kinésine-1 et dynéine localisées directement au niveau de la membrane du RE,
- les MCS du RE avec les endosomes précoces et tardifs, les lysosomes et les mitochondries qui se déplacent le long des microtubules (Visualizing Intracellular Organelle and Cytoskeletal Interactions at Nanoscale Resolution on Millisecond Timescales 2018).
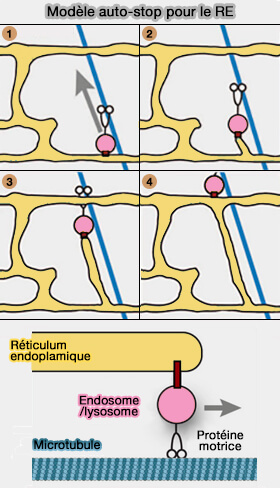
(Figure : vetopsy.fr d'après Sitz et coll)
2. Le déplacement des organites qui provoque la croissance des microtubules est appelé " auto-stop " (hitchhiking en anglais), i.e. un organite fournit le moteur et entraîne le mouvement d'un autre cargo qui n'est pas mobile par lui-même (Peroxisomes, lipid droplets, and endoplasmic reticulum “hitchhike” on motile early endosomes 2015 et Hitchhiking: A Non-Canonical Mode of Microtubule-Based Transport 2017).
3. Des travaux récents suggèrent que ( dynamique du réticulum endoplasmique) :
dynamique du réticulum endoplasmique) :
- 30 à 50% des extensions des tubules du RE sont provoquérs par l'auto-stop sur les endosomes tardifs/lysosomes se déplaçant le long des microtubules,
- 40% des tubules se sont déplacés le long des microtubules indépendamment,
- le reste étant subordonné aux TAC (Tip Attachment complex) ou aux dTAC.
Dynamique du RE et
endosomes précoces
1. La moitié des endosomes positifs pour Rab5, i.e. endosomes précoces, dans les cellules Cos-7 sont attachés au RE (Endoplasmic reticulum–endosome contact increases as endosomes traffic and mature 2013).
- Ils semblent moins mobiles et moins susceptibles de provoquer la mobilité du tubule que les lysosomes associés au RE.
- Toutefois, les endosomes en mouvement peuvent se déplacer vers un tubule du réticulum endoplasmique, le saisir et continuer à se déplacer, tirant le tubule RE derrière eux (Local Cytoskeletal and Organelle Interactions Impact Molecular-Motor-Driven Early Endosomal Trafficking 2013).
(Figure : vetopsy.fr d'après Khattern et coll)
2. Comme les endosomes précoces se déplacent principalement vers le centre cellulaire entraînés par la dynéine ( trafic rétrograde ou centripète des endosomes précoces), l'auto-stop sur les endosomes précoces peut expliquer certaines des extensions des tubules du RE dépendantes de la dynéine (Role of kinesin-1 and cytoplasmic dynein in endoplasmic reticulum movement in VERO cells 2009).
Dynamique du RE et endosomes tardifs/lysosomes
Vue d'ensemble
1. Les lysosomes sont associés et déplacés avec le réseau du RE (The structure and global distribution of the endoplasmic reticulum network are actively regulated by lysosomes 2020).
(Figure : vetopsy.fr d'après Cabukusta et Neefjes)
- Les lysosomes avant, pendant et après un événement d'auto-stop ralentissent lorsqu'ils étaient attachés aux tubules du RE, i.e. la force générée par les moteurs lysosomaux étant amoindrie par le déplacement du tubule (Mobile late endosomes modulate peripheral endoplasmic reticulum network architecture 2021).
- Deux protéines résidentes du RE impliquées dans la synthèse des phospholipides, PIS (PI synthase) et CEPT1 (choline/ethanolamine phosphotransferase 1), se trouvent à l'extrémité des tubules mobiles avec Rab10 (Rab10 GTPase regulates ER dynamics and morphology 2013).
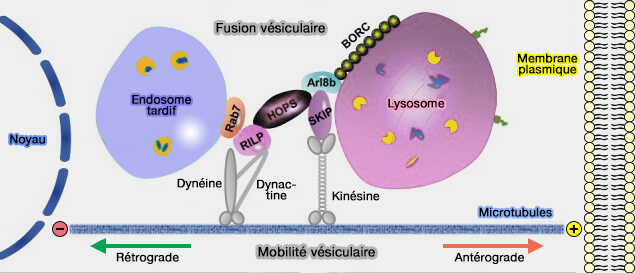
2. La kinésine-1 et la dynéine sont les principaux moteurs du mouvement des endosomes tardifs/lysosomes, et il existe de multiples façons de les recruter et de les contrôler, dont certaines impliquent des interactions avec le RE (transport antérograde et transport rétrograde).

Pour plus de détails, vous pouvez lire : Mechanisms of lysosomal positioning and movement (2018) et Lysosome trafficking and signaling in health and neurodegenerative diseases (2019).
3. Cet auto-stop est crucial pour l'organisation globale du RE.
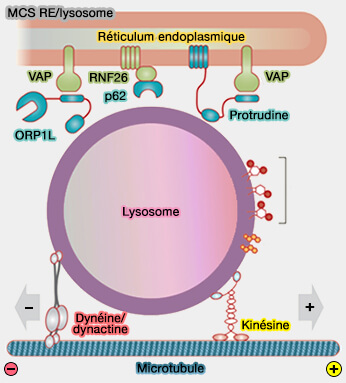
- La perturbation des MCS endosomes tardifs/RE par l'épuisement médié par l'ARNsi de VAPA, qui lie ORP1L pour ancrer l'endosome tardif au RE via Rab7, a réduit l'étendue des tubules du RE et la complexité du réseau.
- L'épuisement de SKIP/PLEKHM2 ou d'Arl8 provoque des changements majeurs dans le réseau, suggérant qu'il s'agit de la principale voie de recrutement de la kinésine-1 pour ce processus, plutôt que de la protrudine/FYCO1.
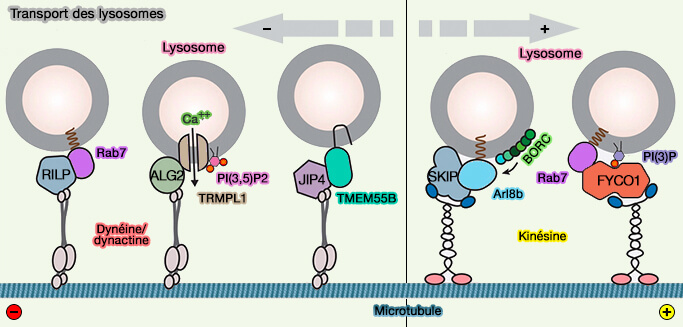
(Figure : vetopsy.fr d'après Cabukusta et Neefjes)
Molécules impliquées
Protrudine
1. La protrudine (produit du gène ZFYVE27) est une protéine transmembranaire endoplasmique avec de multiples partenaires aussi bien au niveau du réticulum endoplasmique (RE) que des endosomes tardifs (Lipid Transfer–Dependent Endosome Maturation Mediated by Protrudin and PDZD8 in Neurons 2020).
Au niveau du RE, elle se lie :
- aux protéines de mise en forme du RE, l'atlastine, les REEP (Receptor Expression-Enhancing Protein) 1 et 5, les réticulons 1, 3 et 4,
- au VAPA, via un motif FFAT.
Au niveau des endosomes tardifs, elle se lie :
- à Rab7-GTP via son domaine de liaison Rab (RBD),
- au PI(3)P via son domaine FYVE.
Ces interactions permettent à la protrudine de relier les membranes du RE et de l'endosome.
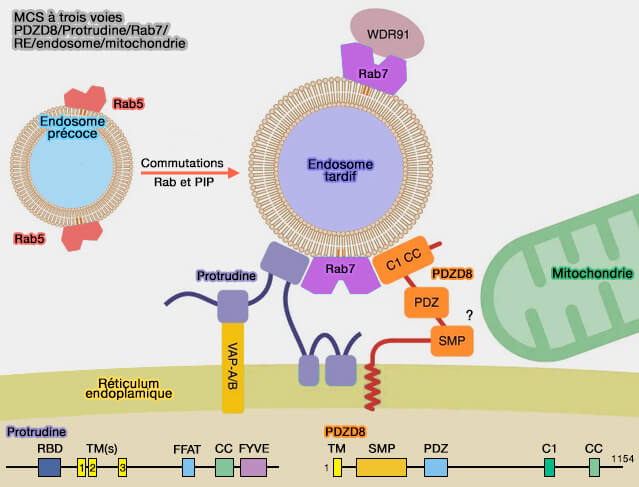
(Figure : vetopsy.fr d'après Elbaz-Alon et coll)
2. La protrudine se lie à tous les membres de la famille KIF5, bien que l'interaction soit la plus forte avec KIF5A (Protrudin serves as an adaptor molecule that connects KIF5 and its cargoes in vesicular transport during process formation 2011).
- Leur mutation provoque une paraplégie spastique héréditaire.
- Leur surexpression provoque la formation de saillies dans les cellules non polarisées, d'où le nom de la protrudine, tandis que la déplétion médiée par l'ARNsi a conduit à une expansion des régions en forme de feuille marquées par CLIMP63 dans la périphérie cellulaire (Protrudin binds atlastins and endoplasmic reticulum-shaping proteins and regulates network formation 2013).
FYCO1 et protrudine
1. La structure de FYCO1 comprend (Crystal structure of the FYCO1 RUN domain suggests possible interfaces with small GTPases 2020) :
- un domaine RUN qui se lie aux petites GTPases,
- un domaine CC (Coiled coil) qui se lie avec la kinésine et avec Rab7,
- un domaine FYVE qui se lie à PI(3)P,
- un domaine LIR (LC3-Interacting Region) pour la liaison avec LC3 (ATG8 chez la levure),
- un domaine GOLD dont on ne connaît pas les liaisons.
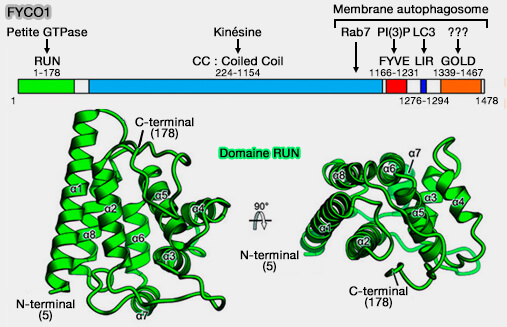
(Figure : vetopsy.fr d'après Sakurai et coll)
2. La protrudine est impliquée dans la formation de MCS avec FYCO1 (FYCO1 is a Rab7 effector that binds to LC3 and PI3P to mediate microtubule plus end-directed vesicle transport 2010).
- La surexpression de la protrudine a conduit à une accumulation d'endosomes tardifs/lysosomes à la périphérie cellulaire, un phénotype qui avait déjà été observé pour FYCO1, une autre protéine liant PI(3)P et Rab7.
-
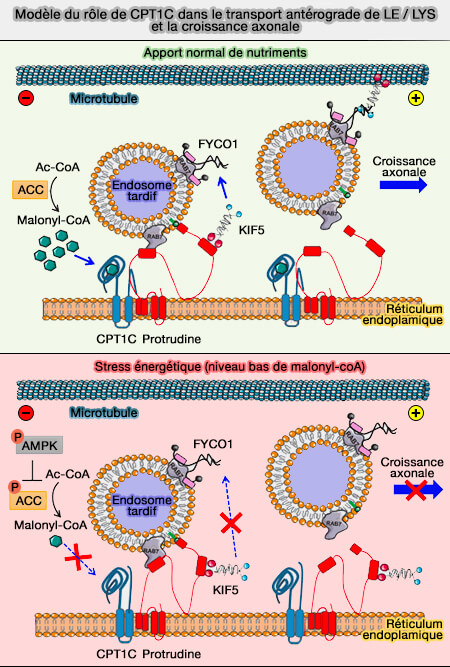
Modèle du rôle de CPT1C
(Figure : vetopsy.fr d'après Palomo-Guerrerro et coll)
3. La kinésine serait transmise de la protrudine à FYCO1 sur l'endosome tardif lors de l'association RE/endosome tardif, activant ainsi le mouvement de l'endosome tardif une fois qu'il s'est libéré du RE.
4. La protrudine et FYCO1 sont impliqués dans :
- l'extension des axones (Axonal transport and neurological disease 2020],
- la formation d'invadopodes, par leur translocation vers la périphérie cellulaire où les endosomes tardifs délivrent la protéase matricielle MT1-MMP (Membrane Type 1P-Matrix Metalloprotease) pour la sécrétion, qui est nécessaire à la migration des cellules cancéreuses (Protrudin-mediated ER–endosome contact sites promote MT1-MMP exocytosis and cell invasion 2020).
5. La position de l'endosome tardif/lysosome est contrôlée par les statuts nutritionnels ( positionnement des lysosomes régulé par les nutriments) et (transport rétrograde ou centripète des endosomes tardifs/lysosomes).
Les taux de cholestérol et d'acides aminés régulent le recrutement ou l'activité de la dynéine et de la kinésine-1.
Cette régulation a un impact majeur sur la dynamique et la distribution des RE au sein de la cellule. Par exemple, la carnitine palmitoyltransférase 1 (CPT1) est une protéine du réticulum régulée par les niveaux de malonyl-CoA (Sensing of nutrients by CPT1C regulates late endosome/lysosome anterograde transport and axon growth 2019).
- Elle permet une croissance neuronale appropriée et contrôle le transport des endosomes/lysosomes tardifs vers la pointe de l'axone, ce qui nécessite sa capacité à se lier au malonyl-CoA.
- En présence de malonyl-CoA, CPT1C favorise le transfert de la kinésine-1 de la protrudine vers FYCO1 sur les endosomes/lysosomes tardifs, favorisant ainsi leur mouvement vers l'extérieur dans les neurones.
Protrudine et PDZD8
1. La protrudine et PDZD8 favorisent de manière coopérative la maturation des endosomes en assurant la médiation (Protrudin and PDZD8 contribute to neuronal integrity by promoting lipid extraction required for endosome maturation 2020) :
- de l'attache RE/endo-lysosome,
- de l'extraction des lipides au niveau des MCS, maintenant ainsi la polarité et l'intégrité neuronales.
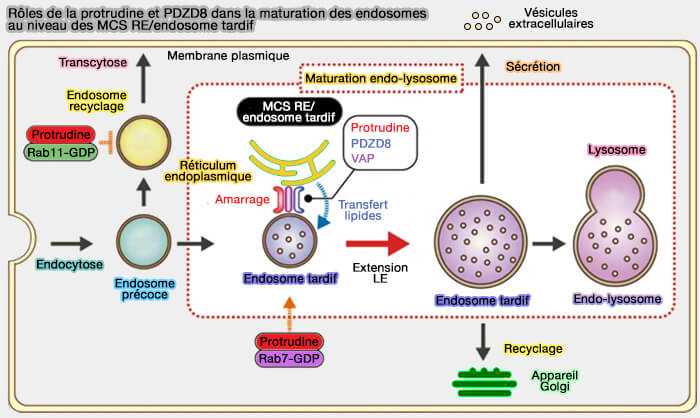
(Figure : vetopsy.fr d'après Cabukusta et Neefjes)
2. PDZD8 partage des similitudes avec Mmm1 de la levure, une sous-unité du complexe ERMES (ER-Mitochondria Encounter Structure), en particulier un domaine SMP (Synaptotagmin-like Mitochondrial lipid-binding Protein) pour le transport des lipides entre le réticulum endoplasmique et les mitochondries chez la levure.
3. PDZD8 jouerait aussi un rôle dans la régulation du Ca++ ( MCS mitochondries/RE et calcium).
Remarque : les lysosomes ont aussi des sites de contact membranaire (MCS) avec :
- les mitochondries,
- l'appareil de Golgi.
Retour aux MCS du réticulum endoplasmique
Biologie cellulaire et moléculaireMembrane plasmiqueNoyauCytoplasmeMitochondriesSystème endomembranaireRéticulum endoplasmiqueAppareil de GolgiEndosomesLysosomesPeroxysomesProtéasomesCytosquelettemicrofilaments d'actineFilaments intermédiairesMicrotubulesMatrice extracellulaireReproduction cellulaireBiochimieTransport membranaireMoteurs moléculairesVoies de signalisation