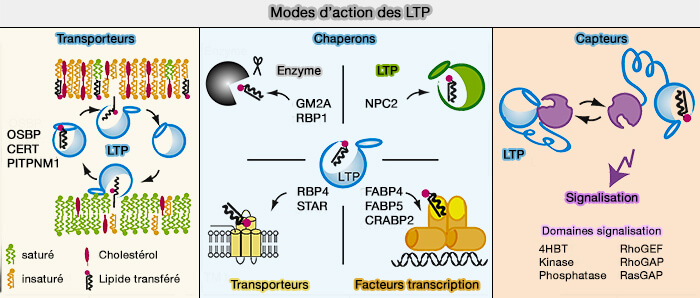
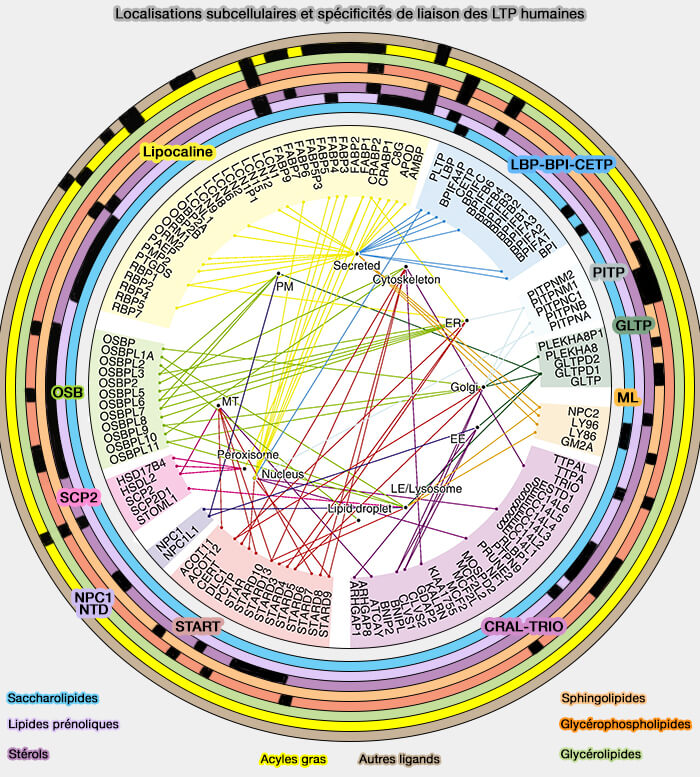
2. Les LTP peuvent remplir au moins l'une des trois fonctions principales dans les cellules en agissant comme ( fonctions des LTP) :
transporteurspour transférer des lipides entre les membranes,
chaperons pour présenter des lipides à une protéine acceptrice, i.e. enzymes, autres LTP, transporteurs transmembranaires ou facteurs de transcription,
capteurs (détecteurs) pour modifier une interaction avec d'autres protéines en réponse à la liaison ou à la libération d'un lipide.
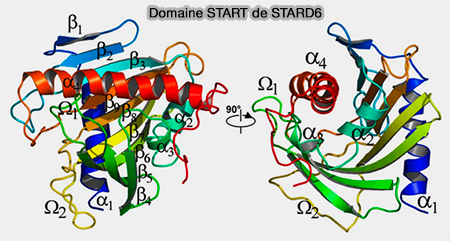
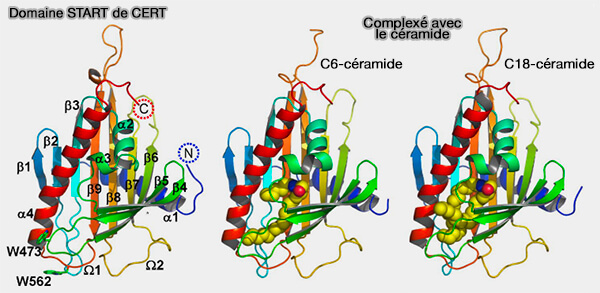
1. Un feuillet β antiparallèle central est ceinturé par les hélices α amino- et carboxy-terminales (α1 et α4), cette dernière étant étroitement refermée sur les 9 brins du feuillet β courbe.
Domaine START de STARD6
(Figure : vetopsy.fr d'après Létourneau et coll)
2. La boucle Ω1 est insérée entre les brins β5 et β6, une autre boucle, Ω2, entre et les brins β7 et β8.
3. La cavité possède une entrée à l’interface du feuillet β et de l’hélice α4, et la partie hydrophobe du lipide est en contact avec la partie hydrophobe de la cavité du domaine. Au fond de la cavité, la partie hydrophile du lipide établit des contacts avec des résidus polaires ou chargés de la protéine.
4. L’entrée d’une molécule de ligand dans le domaine START ne peut se faire que si des réarrangements structuraux majeurs ont lieu ( mécanisme plus bas).
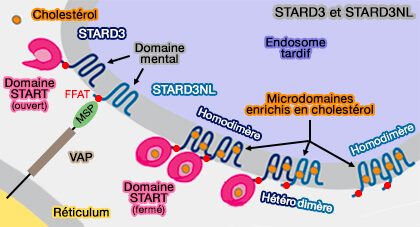
CERT (STARD11 ou COL4A3BP (Collagen type IV alpha-3-binding protein) est étudiée dans un chapitre spécifique.
d. La sous-famille SAM-RhoGAP-START serait impliquée dans la signalisation des petites GTPases Rho et comprend STARD8, STARD12 (ou DCL-1, Deleted in Liver Cancer 1) et STARD13 (ou DLC-2).
e. La sous-famille à activité thioestérase (acyl-CoA thioestérases) comprend STARD14 (ou ACOT11, Acyl-Coenzyme A Thioestérase 1) et STARD15 ou (ou ACOT12)
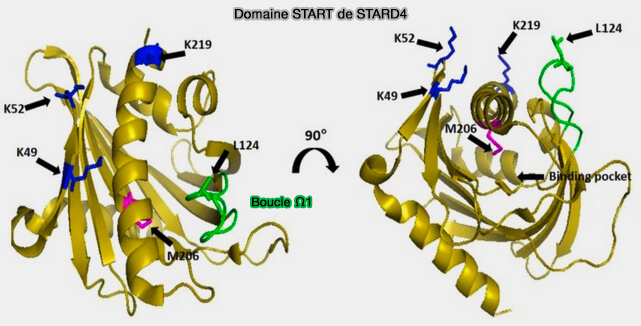
1. STARD4 interagit avec les membranes anioniques à travers un patch basique exposé en surface, i.e. K49, K52 et K219.
Leur mutation diminue le transfert du stérol.
Ces résidus orienterait la protéine dans une position optimale pour le transfert du stérol.
Domaine START de STARD4
(Figure : vetopsy.fr d'après Iaea et coll)
2. La mutation (L124D) dans la boucle Oméga-1 (Ω1), qui recouvre la poche de liaison des stérols, atténue l'activité de transfert des stérols de 100 fois, en réduisant la flexibilité conformationnelle de la protéine, entraînant une diminution du niveau d'interaction membranaire et de transfert de stérol.
3. L'hélice α C-terminale, et non la boucle Ω1, subit une rotation et s'insère dans la bicouche membranaire, ce qui faciliteraient l'exposition de la poche de liaison au stérol par l'ouverture de Ω1p our le transfert du cholestérol vers ou depuis la bicouche lipidique.
Le mécanisme implique donc les mouvements des deux hélices α C-terminale et boucle Ω1.
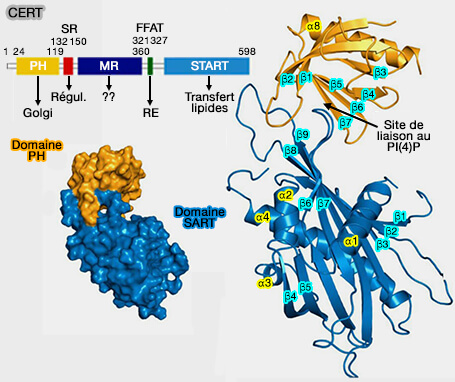
Domaine START de CERT (STARD11)
(Figure : vetopsy.fr d'après Kudo et coll)




 fonctions des LTP) :
fonctions des LTP) :