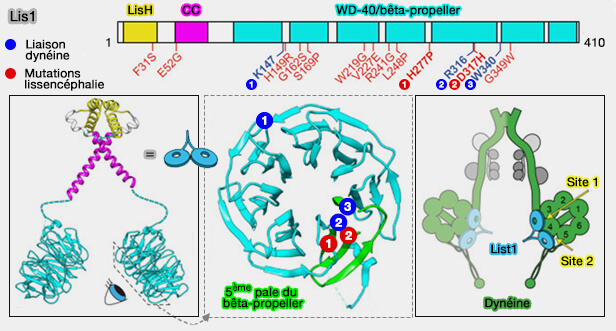
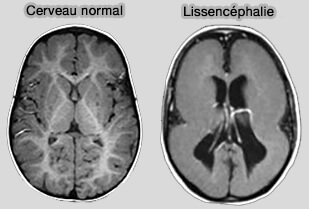
Lissencéphalie
l'initiation du transport du cargo,
le soutien de la capacité de la dynéine à transporter des cargos de forte charge…
Remarque : Lis1 est le principal gène responsable de la lissencéphalie (" cerveau lisse "), maladie cérébrale grave associée à de nombreux symptômes dont des troubles cognitifs et une faible espérance de vie. Lis1 joue un rôle important dans le mouvement des noyaux neuronaux le long des microtubules et une bonne migration neuronale est essentielle pendant le développement du cerveau fœtal.
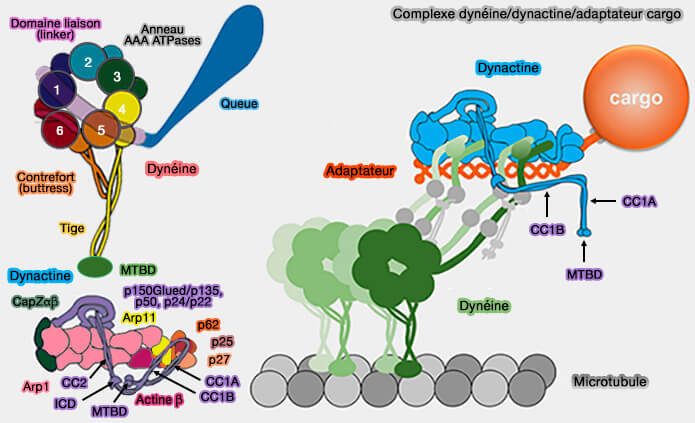
Structure et liaisons
LIS1, qui se présente sous forme dimérique, possède deux domaines fonctionnels distincts .
2. Le grand domaine C-terminal est composé de sept répétitions WD-40, qui sont typiques des structures d'hélices bêta à sept pales (seven-bladed beta-propeller).
Les domaines WD-40 fonctionnent souvent comme des plates-formes d'interaction multi-protéines, ce qui suggère que cette surface pourrait servir d'échafaudage pour le recrutement de divers régulateurs et cibles interagissant avec LIS1 (WD-repeat domain proteins: a novel target class? 2017).
Le mécanisme moléculaire de la régulation médiée par LIS1 paraît complexe car elle peut augmenter ou diminuer la vitesse du déplacement de la dynéine selon les expériences.
Les moteurs de dynéine individuels liés à LIS1 restent attachés aux microtubules pendant de longues périodes, même pendant les cycles d'hydrolyse de l'ATP qui induisent canoniquement le détachement.
Lis1 fonctionne comme un " clutch " (embrayage) qui empêche le domaine AAA1 de la dynéine de transmettre un signal de détachement du microtubule.
Le cycle mécanochimique ne pourrait pas se dérouler serait et verrouillerait d'une manière ou d'une autre l'anneau AAA+ dans un état conformationnel qui conduit à une affinité élevée pour les microtubules (Review: Structure and mechanism of the dynein motor ATPase 2016).
Ce qui paraît curieux car l'affinité élevée pour les microtubules est la configuration droite, i.e. lorsque le domaine de liaison est lié à AAA5
.
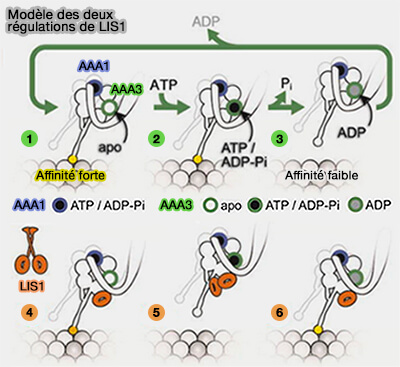
AAA3 pourrait se lier à l'ATP, mais pas l'hydrolyser (4 et 6)
Un deuxième domaine WD-40 de Pac1 pourrait se lier près de la tige du domaine moteur de la dynéine (site 2 et état 5).
Le stade de liaison aux nucléotides du module AAA3 pourrait réguler celui de AA1, site principal d'hydrolyse de l'ATP, à l'origine des changements conformationnels de tout le domaine moteur de la dynéine ( liaison de l'ATP et changements conformationnels).
LIS1 est un activateur de la motilité processive de la dynéine : ce modèle est, semble-t-il, le plus abouti.
Les expériences précédentes ont soulevé la question de savoir comment la liaison de LIS1 à l'anneau AAA+ d'une manière qui stabilise potentiellement un état d'affinité élevée pour les microtubules pourrait augmenter la vitesse de DDA ?
LIS1 favorise probablement l'activité de la dynéine par au moins deux mécanismes distincts.
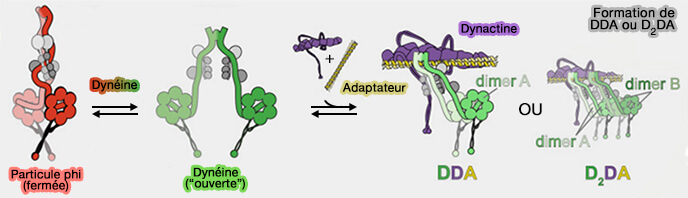
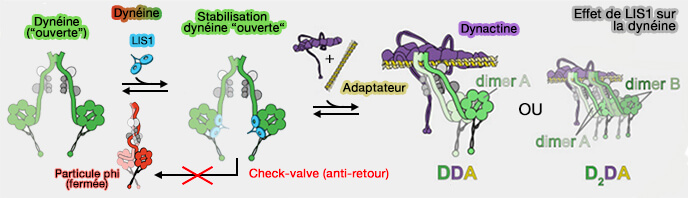
Formation de complexes DDA ou D2DA
(Figure : vetopsy.fr d'après Markus et coll)
1. L'ajout de LIS1 augmente la proportion de complexes DDA assemblés et augmente la vitesse et la force de ces complexes en augmentant la fraction de complexes D2DA.
Les deux dimères de dynéine doivent posséder des domaines moteurs intacts, car l'ajout de complexes de queue de dynéine tronquée a abrogé l'augmentation de vitesse médiée par LIS1.
LIS1 favorise l'assemblage complexe D2DA par un mécanisme qui implique la stabilisation de la conformation " ouverte " de la dynéine, même lorsque on utilise des dynéines mutantes.
2. LIS1 favoriserait aussi l'assemblage du complexe D2DA sans stabilisation de la conformation " ouverte " sans que le mécanisme soit encore connu.
Soit, LIS1 favoriserait directement la liaison de la dynéine à la dynactine (c'est-à-dire par un mécanisme allostérique).
Soit LIS1 n'affecterait que la liaison du deuxième dimère de dynéine, i.e. le dimère de dynéine A lié à LIS1 recrutant le dimère de dynéine B sur la dynactine, en agissant comme un pont entre ces deux dimères de dynéine.
Ce dernier mécanisme est distinct du rôle de LIS1 dans la stabilisation de l'état ouvert, puisqu'un fragment LIS1 monomérique - qui ne possède qu'un seul site de liaison à la dynéine - est suffisant pour favoriser l'assemblage du complexe DDA/D2DA.
Pac1 et LIS1 se lient à la dynéine avec une affinité plus élevée lorsque cette dernière était dans la conformation " ouverte "
LIS1 pourrait être un une molécule de liaison opportuniste : lors du passage stochastique de la dynéine dans la conformation " ouverte ", la liaison de LIS1 à la dynéine empêche son retour dans la conformation phi.
Effet de LIS1 sur la dynéine d'après le modèle " catalytic check valve "
(Figure : vetopsy.fr d'après Markus et coll)
2. Dans ce nouveau modèle, LIS1 agit comme un " catalytic check valve " (clapet anti-retour catalytique), agissant à la fois :
comme un catalyseur pour l'assemblage des complexes DDA et D2DA,
La dissociation de LIS1 du complexe D2DA assemblé fournit une molécule LIS1 de sorte qu'elle pourrait éventuellement agir à nouveau dans un autre cycle d'assemblage D2DA.
La liaison de l'adaptateur pourrait jouer un rôle dans le déclenchement de la dissociation de LIS1 de la dynéine.
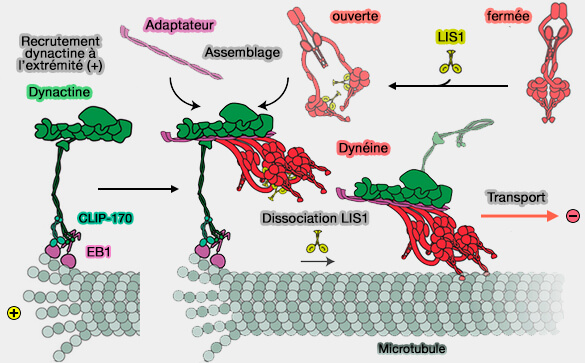
Les molécules de dynéine liées à Pac1, qui s'associent aux extrémités positives via Bik1/CLIP-170 sont dans une configuration " ouverte ", et donc prêtes à l'interaction avec la dynactine.
Ainsi, toute condition dans laquelle l'interaction dynéine-dynactine est renforcée, par exemple, par l'ajout de LIS1, favorisera naturellement la l'association de la dynéine à l'extrémité (+).
La déplétion de LIS1, i.e. mutations ou perte de fonction, doit réduire l'assemblage du complexe dynéine-dynactine et, par conséquent, compromettre la localisation et le transport des cargos.
Remarque : Comment concilier l'activité inhibitrice apparente de Pac1/LIS1 décrite dans le modèle " clutch " avec ces nouvelles données ?
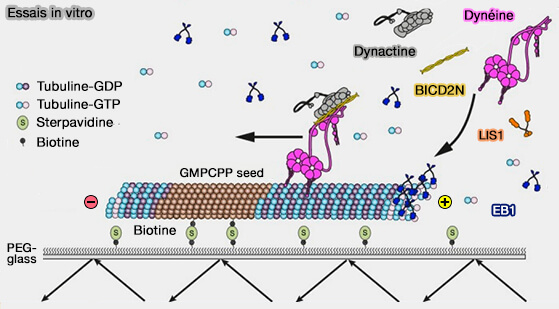
Dans les conditions expérimentales in vitro utilisées pour les études précédentes, Pac1 se lie de manière robuste aux microtubules. Or, in vivo, cela ne semble pas être le cas, mais rien n'est encore tranché.
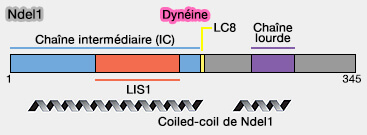
Nde1/Ndel1 ou Nude/Nudel
Les protéines Nde1/Ndel1 (Nuclear Distribution Element 1/Nde-like1), appelées aussi Nude/Nudel, sont des protéines contenant :
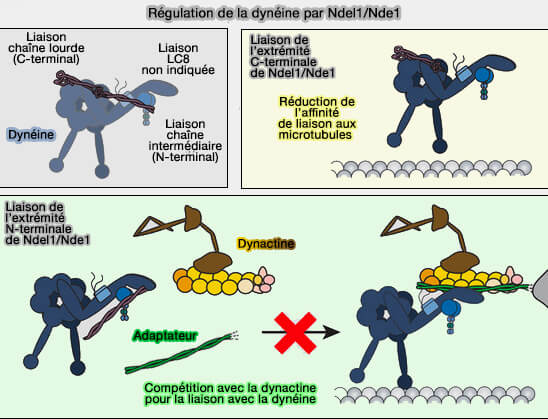
Cette interaction semble innhiber l'interaction avec les microtubules.
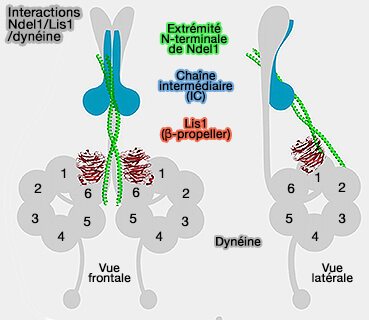
3. Les interactions nécessaires à l'assemblage du complexe tripartite dynéine-LIS1-Nde1/Ndel1 sont mal comprises, et des informations supplémentaires sont nécessaires pour mieux comprendre son organisation, ainsi que la nature et la contribution de chacune de ces interactions.
La liaison compétitive de Nde1/Ndel1 et p150Glued pour DIC suggère un rôle potentiellement supplémentaire pour Nde1/Ndel1 dans la régulation de l'interaction dynéine-dynactine au-delà de la coordination de la liaison dynéine-LIS1
Régulation de la dynéine par Ndel1/Nde1
(Figure : vetopsy.fr d'après Garrott et coll)
Des travaux supplémentaires sont nécessaires pour comprendre la séquence d'interactions qui se produisent pour les nombreux facteurs qui conduisent finalement à l'assemblage DDA/D2
DA.





 modèle " clutch " ou embrayage),
modèle " clutch " ou embrayage),