


Système endo-lysosomal
LRO (Lysosome-Related Organelles)
BLOC-2 et BLOC-3
- Vue d'ensemble du système endomembranaire
- Endosomes
- Lysosomes
- Vue d'ensemble
- Structure des lysosomes
- Fonctions des lysosomes
- Biogenèse des lysosomes
- Biogenèse par livraison des protéines et fusion
- Biogenèse par reformation (fission)
- Biogenèse par régulation transcriptionnelle (TFEB/TFE3/TFEC)
- LRO (Lysosome-Related Organelles)
- Système endo-lysosomal et phosphoinositides
- MCS (sites de contact membranaire) RE/endosomes/lysosomes
- Trafic des endosomes/lysosomes
- Fusion et fission membranaire
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

La biogénèse des LRO (Lysosome-Related Organelles ou organites liées aux lysosomes) nécessite les complexes BLOC (Biogenesis of Lysosome-related Organelles Complex) dont BLOC-2 et BLOC-3.
La biogenèse des LRO (Lysosome-Related Organelles ou organites liées aux lysosomes) nécessite :
- l'adaptine AP-3 et quelquefois AP-1,
- les complexes BLOC (Biogenesis of Lysosome-related Organelles Complex),
- les complexes HOPS et CORVET.

Les adaptines sont étudiées dans un chapitre spécial.
(Figure : vetopsy.fr d'après Flaumenhaft)
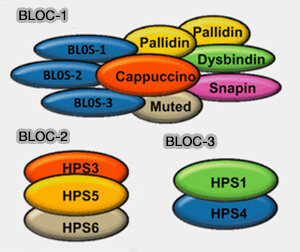
2. On trouve trois complexes BLOC :
Complexe BLOC2

Vous pouvez lire les avancées récentes sur la biogenèse des mélanosomes : Recent advances in understanding the molecular basis of melanogenesis in melanocytes (2020).
1. BLOC-2 est composé de trois grandes sous-unités (Characterization of BLOC-2, a Complex Containing the Hermansky–Pudlak Syndrome Proteins HPS3, HPS5 and HPS6 2004) :
- HPS3, qui contient un domaine longin,
- HPS5,
- HPS6, sous-unité dont les gènes sont mutés dans des variantes HPS correspondantes et des modèles murins de la maladie.
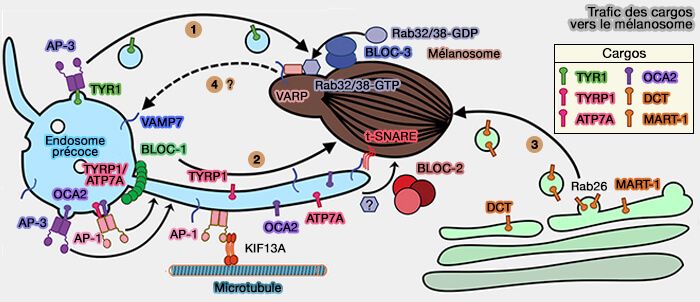
2. La fonction moléculaire de BLOC-2 au cours de la biogenèse LRO n'est pas entièrement connue, mais est mieux comprise dans le contexte de la maturation des mélanosomes (Voie 2).
- Comme BLOC-1, BLOC-2 dans les mélanocytes se localise dans les endosomes tubulaires, , et une cohorte de BLOC-1 et BLOC-2 interagit physiquement, suggérant que BLOC-1 et BLOC-2 fonctionnent dans la même voie (BLOC-1 Interacts with BLOC-2 and the AP-3 Complex to Facilitate Protein Trafficking on Endosomes 2006).
- BLOC-2 influence la livraison mélanosomale des cargos dépendantes de BLOC-1, y compris TYRP1, OCA2, ATP7A et une cohorte de TYR, à partir des endosomes précoces dans les mélanocytes.
(Figure : vetopsy.fr d'après Bowman et coll)
3. BLOC-2 est nécessaire pour que les porteurs tubulaires destinés aux mélanosomes établissent des contacts stables avec les mélanosomes en maturation (BLOC-2 targets recycling endosomal tubules to melanosomes for cargo delivery 2015).
- En l'absence de BLOC-2, ces tubules se forment au même rythme, mais ils ont une durée de vie plus courte, établissent moins de contacts avec les mélanosomes et les contacts qu'ils établissent sont de plus courte durée.
- Les cargos dépendants du BLOC-1 entrent dans les tubules pour le recyclage, mais sont délivrés aux cibles classiques de ces endosomes de recyclage, i.e. l'appareil de Golgi et la membrane plasmique (MPe), entraînant une augmentation du cycle à travers ces organites (BLOC-1 Is Required for Cargo-specific Sorting from Vacuolar Early Endosomes toward Lysosome-related Organelles 2007).
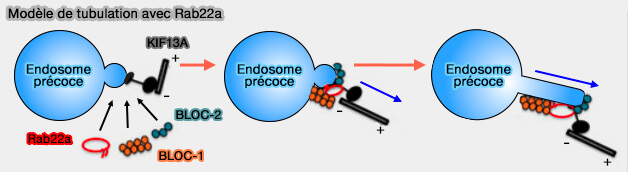
4. BLOC-1 et BLOC-2 jouent avec Rab22a un rôle important dans le système immunitaire ( endosomes de recyclage).
endosomes de recyclage).
(Figure : vetopsy.fr d'après Shakya et coll)
Complexe BLOC3
BLOC-3 et Rab32/38
(Figure : vetopsy.fr d'après Gerondopoulos et coll)
1. BLOC-3 est un complexe à deux sous-unités qui contiennent un domaine longin (Biogenesis of lysosome-related organelles complex 3 (BLOC-3): A complex containing the Hermansky–Pudlak syndrome (HPS) proteins HPS1 and HPS4 2003) :
- HPS1,
- HPS4, à homologie limitée avec les sous-unités MON1 et CCZ1 du facteur d'échange de nucléotides guanine (GEF) pour la petite GTPase Rab7 (Early Origin of Genes Encoding Subunits of Biogenesis of Lysosome-related Organelles Complex-1, -2 and -3 2010).
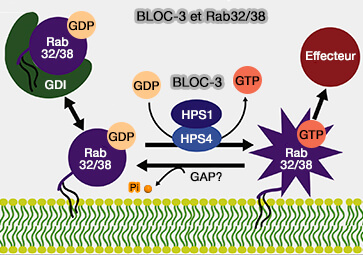
2. BLOC-3 est le principal GEF (Guanine nucleotide exchange factor) pour deux petites GTPases hautement apparentées, Rab32 et Rab38 (BLOC-3 Mutated in Hermansky-Pudlak Syndrome Is a Rab32/38 Guanine Nucleotide Exchange Factor 2012).
a. Rab32/38 ont été impliquées dans la biogenèse :
- des mélanosomes (BLOC-2, AP-3, and AP-1 Proteins Function in Concert with Rab38 and Rab32 Proteins to Mediate Protein Trafficking to Lysosome-related Organelles 2010),
- des granules denses des plaquettes (Mechanism of platelet dense granule biogenesis: study of cargo transport and function of Rab32 and Rab38 in a model system 2012),
- des corps lamellaires AT2 (Rab38 targets to lamellar bodies and normalizes their sizes in lung alveolar type II epithelial cells 2011).
(Figure : vetopsy.fr d'après Bowman et coll)
b. Rab32 et Rab38 se localisent en grande partie dans les mélanosomes, ainsi que dans les structures tubulo-vésiculaires à proximité des mélanosomes et de l'appareil de Golgi.
Leur épuisement conduit à une mauvaise localisation des cargos de mélanosomes telles que TYRP1 et TYR (Rab38 and Rab32 control post-Golgi trafficking of melanogenic enzyme 2006).
(Figure : vetopsy.fr d'après Dennis et coll)
Remarque : Rab32/38 sont impliquées dans le trafic du transporteur de glucose GLUT1 vers la membrane plasmique en régulant la molécule effectrice VARP.
Rôles de BLOC-3
et Rab32/38
Transport endosomal
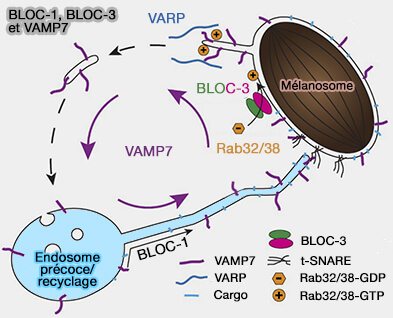
1. BLOC-3 et Rab32/38 semblent jouer un rôle primordial dans le transport rétrograde (centripète) à partir des mélanosomes (BLOC-1 and BLOC-3 regulate VAMP7 cycling to and from melanosomes via distinct tubular transport carriers 2016).
- Les mélanosomes émettent de courts tubules mobiles qui sont enrichis en VAMP7 (ou SYBL1), une protéine de fusion SNARE vésiculaire nécessaire au trafic de fret antérograde dépendant de BLOC-1 (voie 4).
- Les tubules sont également enrichis en Rab38 et son effecteur, VARP, une protéine d'échafaudage qui soutient probablement l'incorporation de VAMP7 dans ceux-ci (Structure-Function Analysis of VPS9-Ankyrin-repeat Protein (Varp) in the Trafficking of Tyrosinase-related Protein 1 in Melanocytes 2011).
(Figure : vetopsy.fr d'après Ripoll et coll)
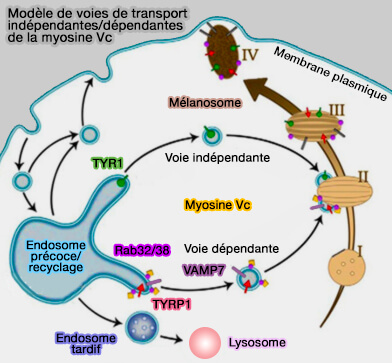
2. Un autre effecteur des Rab32/38 est la myosine Vc, dont l'inactivation a également un impact sur le trafic antérograde (centrifuge) des cargos de mélanosomes (Myosin Vc Interacts with Rab32 and Rab38 Proteins and Works in the Biogenesis and Secretion of Melanosomes 2014).
Fission
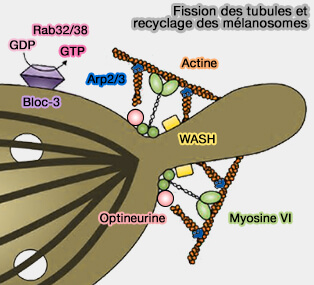
1. BLOC-3 et Rab32/38 provoquent aussi la fission des tubules intermédiaires qui émergent des mélanosomes pour recycler les membranes et les protéines grâce aux protéines SNARE (Myosin VI and branched actin filaments mediate membrane constriction and fission of melanosomal tubule carriers 2018).
- Le complexe WASH active localement la nucléation du réseau d'actine ramifié par l'ARP2/3 sur les tubules pour commencer à former le collet d'actomyosine pour la fission de tubule.
- La zone de scission des tubules, délimitée par l'optineurine, une protéine adaptatrice recrutée également au cou des tubules, recrute la myosine VI, myosine atypique qui se déplace vers l'extrémité moins du microfiament d'actine-F, contrairement aux autres myosines.
2. Toutefois, un nouveau mécanisme de fission membranaire appelé friction-driven scission (FDS) s'appuie sur les forces de friction générées par des protéines localisées au cou des vésicules ou des tubules intermédiaires, qui peuvent alors effectuer leur fission (Friction Mediates Scission of Tubular Membranes Scaffolded by BAR Proteins 2017).
Le modèle FDS est étudié dans un chapitre spécial.
(Figure : vetopsy.fr d'après Bultema et coll)
a. La myosine VI permet de croiser l'actine au cou pour produire la force nécessaire pour resserrer le cou et promouvoir la fission et la libération du tubule.
- Finalement, les protéines à domaine BAR, les moteurs et l'actine se désassemblent de la vésicule endocytée, qui continue le long de la voie endocytaire.
- Les tubules libérés recyclent Vamp7 pour une réutilisation possible, vraisemblablement vers les endosomes précoces, et probablement exportent également des composants supplémentaires pour permettre une autre maturation mélanosomique.
b. La myosine VI a aussi un rôle dans :
- le transport des cargos dans les endosomes initiaux ( GAIP et myosine VI),
- le détachement de la membrane des vésicules de clathrine (rôles des myosines dans les stades 4 et 5 de l'endocytose clathrine-dépendante).
Autres rôles
BLOC-3 et Rab32/38 ont aussi un rôle dans :
- le fonctionnement pulmonaire,
- l'inflammation.
Retour aux LRO
Biologie cellulaire et moléculaireMembrane plasmiqueNoyauCytoplasmeMitochondriesSystème endomembranaireRéticulum endoplasmiqueAppareil de GolgiEndosomesLysosomesPeroxysomesProtéasomesCytosquelettemicrofilaments d'actineFilaments intermédiairesMicrotubulesMatrice extracellulaireReproduction cellulaireBiochimieTransport membranaireMoteurs moléculairesVoies de signalisation