


Système endo-lysosomal
Trafic des endosomes/lysosomes
Trafic lié au cytosquelette d'actine
- Vue d'ensemble du système endomembranaire
- Endosomes
- Lysosomes
- Trafic des endosomes/lysosomes
- Fusion et fission membranaire
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

La distribution spatio-temporelle des organites endo-lysosomaux dépend du transport entraîné par des moteurs de microtubules tels que les kinésines et la dynéine, et par des moteurs de myosine à base d'actine.
Les organites se déplacent dans tout le cytoplasme le long du cytosquelette, i.e. microtubules et filaments d'actine, et ces mouvements sont finement régulés ( vue d'ensemble du trafic des endosomes).
vue d'ensemble du trafic des endosomes).
1. Les protéines motrices de microtubules transportent les organites rapidement sur de grandes distances et de manière bidirectionnelle.
- La plupart des kinésines déplacent les organites de l'extrémité moins vers l'extrémité plus (transport antérograde ou centrifuge).
- La dynéine provoque le déplacement en sens inverse, de l'extrémité plus vers l'extrémité moins (transport rétrograde ou centripète).
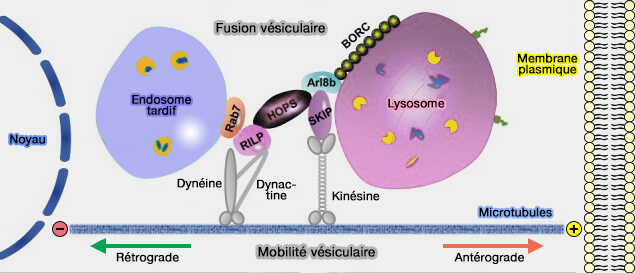
(Figure : vetopsy.fr d'après Khattern et coll)
2. Les moteurs de l'actine, i.e. les myosines, interviennent sur de courtes distances et de manière lente.

Vous pouvez lire : F-Actin Dynamics in the Regulation of Endosomal Recycling and Immune Synapse Assembly (2021).
Vue d'ensemble du transport par le cytosquelette d'actine
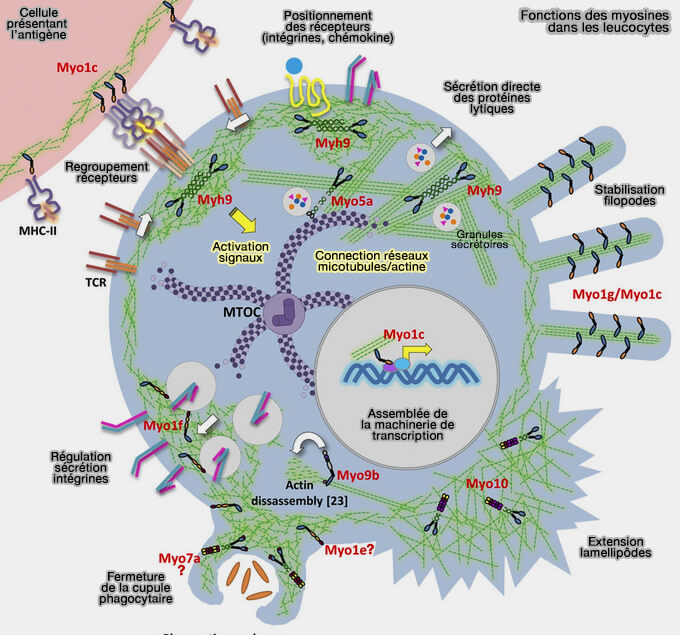
1. Le cytosquelette d'actine et les myosines, protéines motrices, une trentaine environ avec différentes fonctions et modèles d'expression, modulent également le transport des organites endo-lysosomaux par une variété de mécanismes.
Ce transport a lieu sur de courtes distances par rapport à celui des microtubules.

Les myosines sont étudiées dans un chapitre spécial avec les moteurs moléculaires.
2. Ce mécanisme correspond également à un modèle petite GTPase-adaptateur-moteur similaire à celui observé pour les interactions des organites avec les moteurs des microtubules.
Cortex cellulaire
L'un de ces mécanismes implique la capture dynamique d'organites dans les régions cellulaires riches en actine telles que le cortex d'acto-myosine qui sous-tend la membrane plasmique ( actine et cortex cellulaire).
Le cortex d'acto-myosine cellulaire est étudié dans un chapitre spécial.
(Figure : vetopsy.fr d'après Scklnick et coll)
Endosomes et lysosomes
1. Le positionnement et la fusion des lysosomes sont sous le contrôle de moteurs de myosine coordonnés par de petites GTPases Rab ( lysosomes et membrane plasmique).
Par exemple, Rab3a et Rab10 sont impliquées dans la réparation membranaire.
2. Les petites GTPases Rab, en particulier Rab27, sont aussi impliquées dans le trafic des endosomes ou corps multivésiculaires (MVE/MVB) vers la membrane plasmique et leur amarrage pour la libération d’exosomes (trafic et libération des exosomes)
LRO
Pour des LRO (Lysosome-Related Organelles), le mécanisme fait intervenir généralement des complexes composés de :
- de Rab27a,
- de protéines adaptatrices,
- de la myosine Va, et parfois VIIa, pour la rétention périphérique et/ou l'exocytose du contenu du LRO.
(Figure : vetopsy.fr d'après Paul et coll)
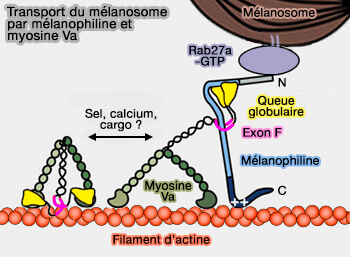
Mélanosomes
1. Pour les mélanosomes, Rab27A recrute (Rab27 Effectors, Pleiotropic Regulators in Secretory Pathways 2013) :
- la myosine Va et myosine VIIa, via son effecteur Slp2, pour accumuler les mélanosomes à la périphérie cellulaire, sous la membrane plasmique des mélanocytes (Melanosomes on the move: a model to understand organelle dynamics 2011),
- la myosine Va par la mélanophiline (A Coiled-Coil Domain of Melanophilin Is Essential for Myosin Va Recruitment and Melanosome Transport in Melanocytes 2006 et More Than Just a Cargo Adapter, Melanophilin Prolongs and Slows Processive Runs of Myosin Va 2013).
Les mutations de Rab27A provoquent le syndrome de Griscelli de type 2 par défaut de sécrétion des mélanosomes.
2. Pour les autres LRO, Rab27A recrute :
- Munc13-4 pour les granules lytiques des lymphocytes T cytotoxiques (Rab27a Is Required for Regulated Secretion in Cytotoxic T Lymphocytes 2001et Munc13-4 Is an Effector of Rab27a and Controls Secretion of Lysosomes in Hematopoietic Cells 2005).
- Slac2c/MyRIP pour les granules de sécrétion d'insuline dans les cellules β pancréatiques (Involvement of the Rab27 Binding Protein Slac2c/MyRIP in Insulin Exocytosis 2003).
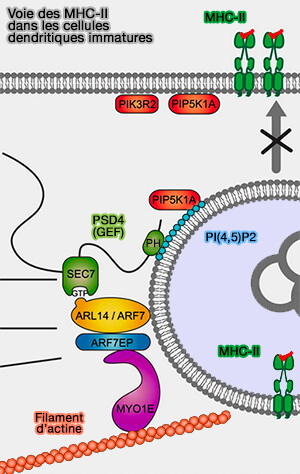
3. Les CMH de classe II (MIIC) sont des LRO qui fusionnent avec la membrane plasmique lors de la maturation des cellules dendritiques immatures.
Dans ces cellules, les MIIC sont maintenus dans la cellule par l'action de (A Genome-wide Multidimensional RNAi Screen Reveals Pathways Controlling MHC Class II Antigen Presentation 2011) :
- la petite GTPase Arl14,
- l'effecteur ARF7EP,
- Myo1e, une myosine à une seule jambe qui est incapable de se déplacer, mais qui peut attacher les cargos à l'actine.
(Figure : vetopsy.fr d'après Maravillas-Montero et coll)
Queues de comète d'actine
Un autre mécanisme de motilité basé sur l'actine implique la formation de queues de comètes d'actine sur les endosomes et les lysosomes.
1. Ces queues :
- les propulsent à travers le cytoplasme,
- contribuent au bourgeonnement et à la fission des tubules endosomaux.
Le complexe WASH et le complexe BLOC-1 jouent un rôle essentiel dans la nucléation de l'actine pour ce type de motilité ( BLOC-1 et microsquelette).
2. L'actine non corticale et les myosines jouent un rôle essentiel dans la biologie des lysosomes.
- La myosine Ia facilite la livraison du matériel endocytosé aux lysosomes (Association of Myosin I Alpha with Endosomes and Lysosomes in Mammalian Cells 1999).
- La myosine Ic est impliquée dans la fusion des autophagosomes avec les lysosomes (Loss of functional MYO1C/myosin 1c, a motor protein involved in lipid raft trafficking, disrupts autophagosome-lysosome fusion 2014).
Cytosquelette d'actine et microtubules

Le cytosquelette d'actine pourrait contribuer à la motilité et au positionnement endo-lysosomal de manière supplémentaire, fonctionnant :
- soit en coopération (Actin Filaments and Myosin I Alpha Cooperate with Microtubules for the Movement of Lysosomes 2001 et BLOC-1 Brings Together the Actin and Microtubule Cytoskeletons to Generate Recycling Endosomes 2016),
- soit en opposition aux microtubules (Active diffusion and microtubule-based transport oppose myosin forces to position organelles in cells 2016).
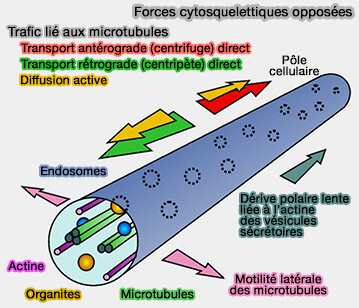
(Figure : vetopsy.fr d'après Lin et coll)
1. Par exemple, les microtubules mélangent et distribuent les peroxysomes.
- Ils soutiennent la motilité bidirectionnelle des endosomes précoces impliqués dans la transport direct centrifuge ou antérograde et rétrograde ou centripète des peroxysomes.
- Ils permettent également leur diffusion active le long de l'axe de la cellule.
2. La flexion des microtubules exerce une force latérale qui augmente le contact entre les peroxysomes et les endosomes précoces.
Ces mécanismes s'opposent à une lente dérive vers le pôle des peroxysomes, médiée par la myosine V à la périphérie cellulaire.
La diffusion thermique ne joue qu'un rôle mineur dans la mobilité des PO et est exclue du diagramme.
Retour à les endosomes
Biologie cellulaire et moléculaireMembrane plasmiqueNoyauCytoplasmeMitochondriesSystème endomembranaireRéticulum endoplasmiqueAppareil de GolgiEndosomes/LysosomesPeroxysomesProtéasomesCytosquelettemicrofilaments d'actineFilaments intermédiairesMicrotubulesMatrice extracellulaireReproduction cellulaireBiochimieTransport membranaireMoteurs moléculairesVoies de signalisation