Le rétromère utilise des protéines accessoires comme les nexines, les Rab et le complexe WASH pour moduler le trafic endocytaire et l'homéostasie mitochondriale.
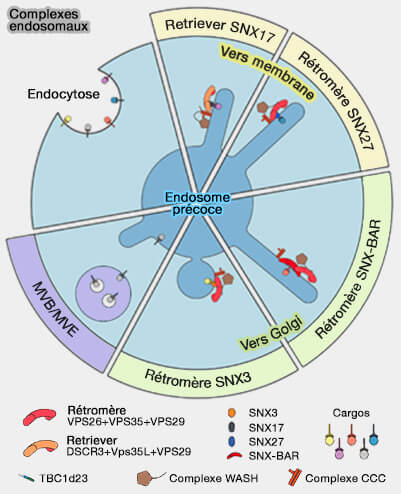
Vue d'ensemble du recrutement du CSC
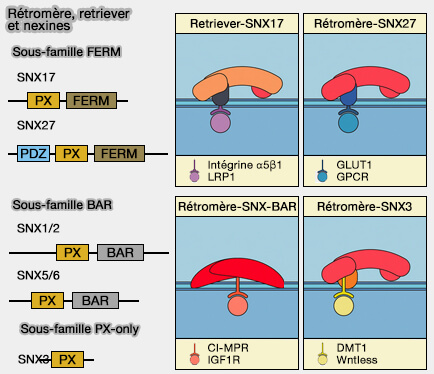
Le rétromère est recruté sur les endosomes par des motifs d'acides aminés courts ou de signaux de tri trouvés dans les protéines membranaires.
Complexes endosomaux
(Figure : vetopsy.fr d'après Wang et all)
Comme le rétromère ne peut se lier directement aux lipides membranaires, il doit utiliser de multiples interactions protéine/protéine.
1. Des adaptateurs de cargos peuvent être associés à la membrane.
2. Le rétromère/SNX3 est formé par le rétromère lié à SNX3, une nexine contenant seulement un domaine PX (sous-famille PX-only), qui, quoique sans domaine BAR, peut provoquer une tubulation in vitro.
La complexité de la biologie des rétromères chez les métazoaires reflète probablement à la fois l'augmentation du nombre de cargos et la diversité des routes de trafic à partir des endosomes.
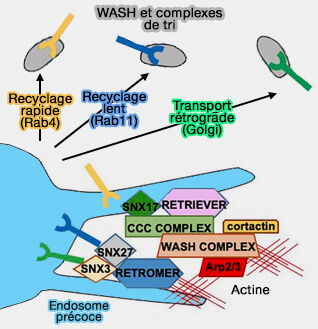
Petites GTPases Rab
Les petites GTPases, essentielles à la formation de vésicules endocytaires enveloppées, aussi bien à base de clathrine, de COPI et de COPII, sont aussi des régulateurs du trafic vésiculaire médié par le rétromère.
2. De tous les régulateurs endogènes connus du rétromère, TBC1D5 affiche la plus grande affinité envers le rétromère (constante de dissociation de 200 nM à 450nM).
Elle est comparable à celle mesurée entre VPS29 et VPS35 (200 nM).
Elle
est supérieure d'au moins un ordre de grandeur à l'affinité entre le rétromère et ses autres partenaires de liaison, tels que SNX3, VARP, Rab7a ou la sous-unité du complexe WASH, FAM21.
Le domaine TBC assure la médiation de l'interaction avec le rétromère, en contactant à la fois VPS35 et VPS2921.
Une boucle de TBC1D5 se lie à une poche hydrophobe conservée sur VPS29, et une seconde boucle peut interagir avec l'extrémité N-terminale de VPS35.
Un mutant VPS35 incapable de s'associer à TBC1D5 ne s'est pas correctement localisé dans les endosomes, ce qui suggère que TBC1D5 pourrait jouer un rôle dans le recrutement de VPS35 dans la membrane endosomale.
L'association étroite entre TBC1D5 et le rétromère s'apparente au complexe Sec23-Sec24 dans le manteau COPII.
La plupart des protéines TBC utilisent un mécanisme catalytique à double doigt arginine-glutamine pour inactiver les substrats Rab, qui est largement différent du mécanisme à doigt arginine unique utilisé par de nombreux GAP des autres petits GTPases.
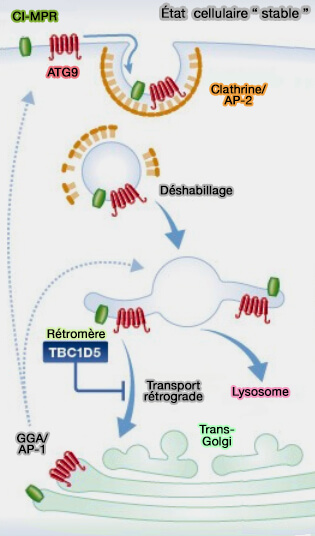
b. En l'absence de rétromère ou de TBC1D5, les niveaux de Rab7a-GTP augmentent considérablement et Rab7a s'accumule sur les domaines lysosomaux, ce qui entraîne :
une diminution de la mobilisation de Rab7a,
un épuisement de Rab7a inactif des autres organites subcellulaires,
un renouvellement membranaire défectueux.
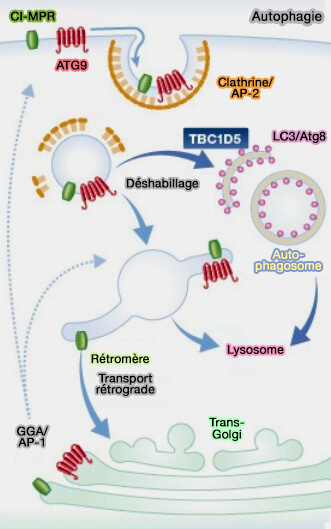
4. TBC1D5 joue également un rôle dans l'autophagie.
Le TBC1D15 mitochondrial semble coexister en équilibre avec son pool cytosolique et Arl8b/SKIP/HOPS puise dans ce dernier pour procéder à l'élimination de Rab7 de certains MVB.




 rétromère/SNX3).
rétromère/SNX3).