


Fission membranaire
Fission membranaire active
- Biologie cellulaire et moléculaire
- Constituants de la cellule
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

Les fissions membranaires actives utilisent de l'énergie selon plusieurs mécanismes.
La fission membranaire, complexe et faisant intervenir plusieurs protéines et les phospholipides, peut être (Diversity of Biological Membrane Fission Mechanisms 2017) :
- passive, sans utilisation d'énergie,
- active.

Vous pouvez avoir un aperçu des fissions membranaires dans un grand nombre de processus avec une bibliographie fournie et récente dans : Protein Amphipathic Helix Insertion: A Mechanism to Induce Membrane Fission (2019).
Les fissions actives peuvent nécessiter :
a. la constriction du cou de la vésicule par :
- des protéines externes, comme la dynamine ou les membres de la superfamille de la dynamine (DSP),
- des protéines internes comme le complexe ESCRT, en particulier ESCRT-III.
b. des réorganisations des domaines lipidiques comme avec les protéines CtBP/BARS,
c. l'appui du cytosquelette :
- filaments d'actine en lien avec les phosphoinositides,
- microtubules par un mécanisme de fission membranaire appelé friction-driven scission (FDS).
Les fissions spécifiques sont le plus souvent traitées dans des chapitres particuliers aux organites.
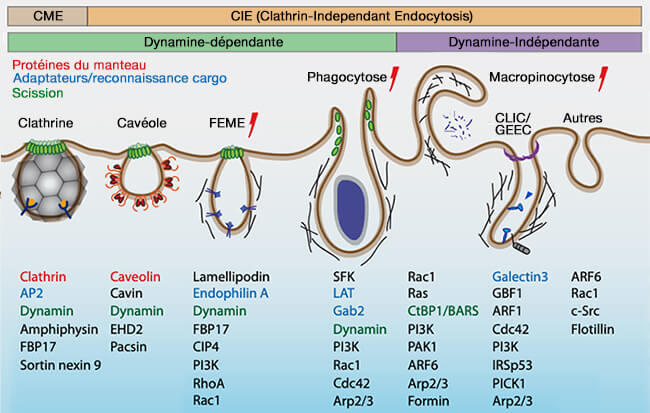
(Figure : vetopsy.fr d'après Thottacherry et coll)
Protéines de fission
Dynamines et DSP
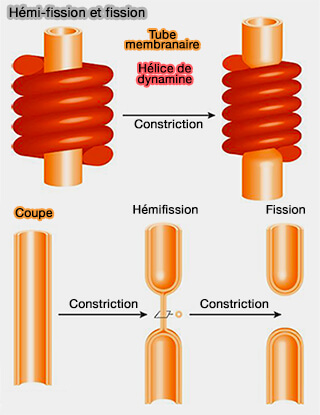
(Figure : vetopsy.fr d'après Antonny et coll)
1. Le mécanisme le plus étudié est celui des dynamines, i.e. GTPases, qui possèdent trois propriétés.
- 1. Elles s'auto-oligomérisent en hélices autour d'un tube membranaire (
 oligomérisation de la dynamine).
oligomérisation de la dynamine). - 2. Les changements conformationnels induits par les nucléotides conduisent à une constriction du polymère et donc de la membrane (dynamine et GPT).
- 3. Les dynamines induisent la fission des cols membranaires par l'hydrolyse du GTP.

La structure de la dynamine est étudiée longuement avec son rôle enzymatique de GTPase et son rôle dans l'endocytose ( détachement de la vésicule).
2. La dynamine, par son domaine PRD C-terminal, entre en interaction avec de nombreux partenaires ( dynamine et partenaires de fission).
a. Les protéines à domaine BAR courbent la membrane comme l'endophiline ou l'amphiphysine.
b. Les protéines à domaine SH3 sont :
- des protéines régulatrices de l'actine,
- des protéines de signalisation,
- des enzymes métabolisant les phosphoinositides membranaires.
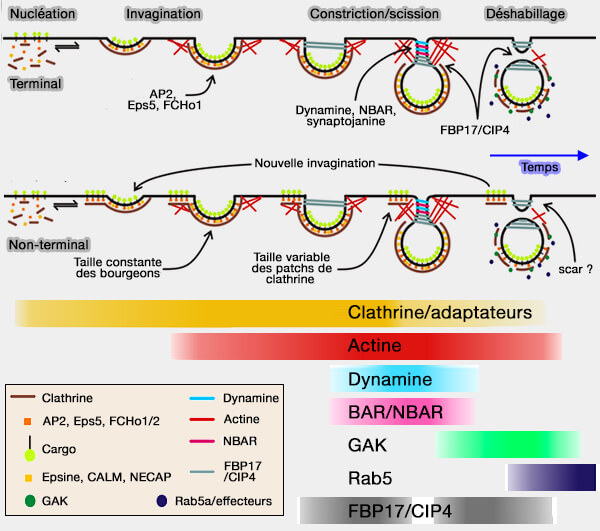
(Figure : vetopsy.fr d'après Taylor et coll)
Remarque : d'autres membres de la superfamille de la dynamine (DSP) interviennent dans la fission mitochondriale comme dans d'autres fissions ( fission mitochondiale).
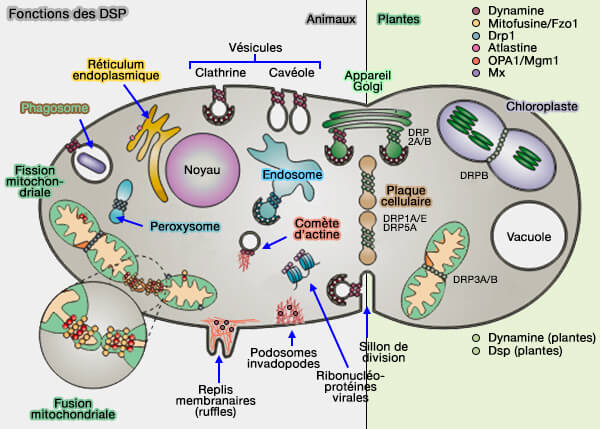
(Figure : vetopsy.fr modifiée d'après Praefcke et McMahon)
Complexe ESCRT
Le complexe ESCRT a plusieurs rôles qui impliquent des processus de remodelage cellulaire, et en particulier des déformations membranaires.
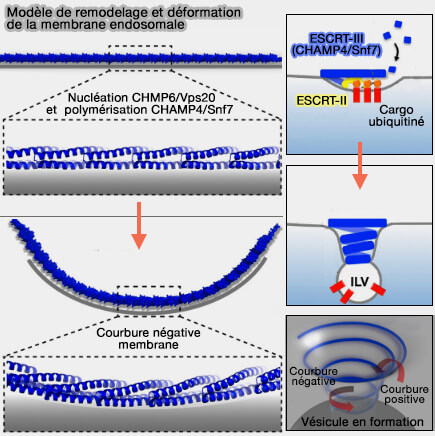
Remodelage et déformation de la membrane endosomale
(Figure : vetopsy.fr d'après Tang
et coll)
1. ESCRT-III est considéré comme le principal moteur du remodelage et de la scission membranaire dans le système endolysosomal.
L'ESCRT-III et Vsp4, ainsi que les différents modèles de scission membranaire, sont étudiés dans des chapitres spéciaux.
2. ESCRT-IIII est impliqué dans :
- la biogenèse des vésicules intraluminales (ILV) et des endosomes ou corps multivésiculaires (MVE/MVB),
- la cytokinèse,
- le bourgeonnement viral.
CtBP/BARS et
réorganisation lipidique
1. Les protéines CtBP/BARS, CtBP1 et CtBP2 (C-terminal-binding protein/Brefeldin A ADP-Ribosylated Substrat) n'ont rien à voir avec les protéines à domaine BAR.
Les protéines CtBP/BARS sont étudiées dans un chapitre spécial.
Dans le cytoplasme, elles entrent en interaction avec les lipides, i.e. PI(4)P dans le réseau trans-Golgi (TGN) ou l'acide phosphatidique (PA) dans le cis-Golgi, sans que l'on comprenne bien les mécanismes sous-jacents.
2. Les CtBP sont impliquées dans la fission membranaire, i.e par :
- la formation de porteurs post-golgiens (PGC) au réseau trans-Golgi (TGN),
- la libération de vésicules COPI au cis-Golgi,
- l'endocytose des fluides au niveau du Golgi et de la membrane plasmique,
- la macropinocytose au niveau de la membrane plasmique.
Remarque : les CtBP interviennent aussi dans la signalisation présynapse/noyau par leur interaction avec Bassoon et Piccolo.
Cytosquelette et fission
Microfilaments d'actine et phosphoinositides
1. Les microfilaments d'actine sont essentiels à l'endocytose, en particulier à l'endocytose clathrine-dépendante ( recrutement de l'actine pendant la CME).
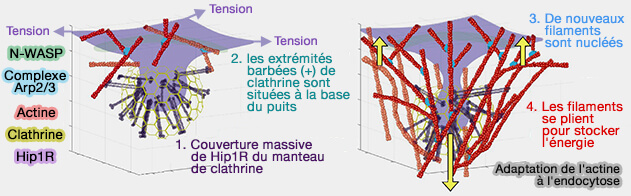
(Figure : vetopsy.fr d'après Akamatsu et coll)
2. La polymérisation de l'actine sur les membranes favoriserait la formation de domaines lipidiques, i.e. PI(4,5)P2 ou PIP2, conduisant à l'apparition d'une tension de ligne ( cf. plus haut), ce qui pourrait contribuer à
la scission (Actin Polymerization Serves as a Membrane Domain Switch in Model Lipid Bilayers 2006 et The Mechanochemistry of Endocytosis 2009).
3. Les myosines, en particulier la myosine VI, interviendraient aussi dans la fission ( rôles des myosines dans l'endocytose).
Microtubules, protéines BAR et modèle FDS
Un mécanisme de fission membranaire appelé friction-driven scission (FDS) s'appuie sur les forces de friction générées par des protéines localisées au cou des vésicules ou des tubules intermédiaires, qui peuvent alors effectuer leur fission (Friction Mediates Scission of Tubular Membranes Scaffolded by BAR Proteins 2017).
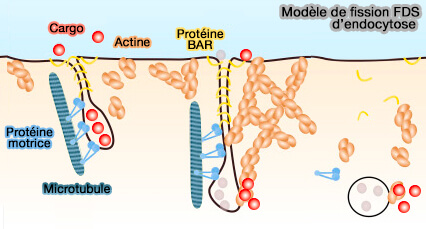
(Figure : vetopsy.fr d'après Simunovic et coll)
Le modèle d'endocytose généré par le mécanisme FDS est le suivant.
- 1. Les protéines à domaine BAR, comme l'endophilline, se lient au feuillet cytosolique des invaginations de la membrane endocytaire, formant un échafaudage.
- 2. Le tube membranaire contenant les cargos est allongé par des moteurs moléculaires des microtubules, tels que les dynéines ou les kinésines.
- 3. L'allongement du tube est facilité par la polymérisation de l'actine, jusqu'au point où le FDS se produit.
- 4. La myosine VI permet de croiser l'actine au cou pour produire la force nécessaire pour resserrer le cou et promouvoir la fission et la libération du tubule.
- 5. Finalement, les protéines du domaine BAR, les moteurs et l'actine se désassemblent de la vésicule endocytée, qui continue son chemin le long de la voie endocytaire.
La myosine VI a aussi un rôle dans le transport des cargos dans les endosomes initiaux et dans la fission des mélanosomes ( GAIP et myosine VI).
Mécanisme de fission
Biologie cellulaire et moléculaireConstituants de la celluleTransport membranaireTransports sans mouvements membranairesTransports passifsTransports actifsTransports avec mouvements membranaires : trafic vésiculaireFusion/fission membranaireEndocytoseVoie sécrétoireCanaux ioniquesTransporteursUniportsPompesCo-transporteursMoteurs moléculairesVoies de signalisation