


Système endo-lysosomal
LRO (Lysosome-Related Organelles)
Complexes BLOC et BLOC-1
- Vue d'ensemble du système endomembranaire
- Endosomes
- Lysosomes
- Vue d'ensemble
- Structure des lysosomes
- Fonctions des lysosomes
- Biogenèse des lysosomes
- Biogenèse par livraison des protéines et fusion
- Biogenèse par reformation (fission)
- Biogenèse par régulation transcriptionnelle (TFEB/TFE3/TFEC)
- LRO (Lysosome-Related Organelles)
- Système endo-lysosomal et phosphoinositides
- MCS (sites de contact membranaire) RE/endosomes/lysosomes
- Trafic des endosomes/lysosomes
- Fusion et fission membranaire
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

La biogénèse des LRO (Lysosome-Related Organelles ou organites liées aux lysosomes) nécessite les complexes BLOC (Biogenesis of Lysosome-related Organelles Complex) dont BLOC-1.
La biogenèse des LRO (Lysosome-Related Organelles ou organites liées aux lysosomes) nécessite :
- l'adaptine AP-3 et quelquefois AP-1,
- les complexes BLOC (Biogenesis of Lysosome-related Organelles Complex),
- les complexes HOPS et CORVET.

Les adaptines, les complexes HOPS et CORVET sont étudiés dans un chapitre spécial.
Vue d'ensemble des complexes BLOC

Ce chapitre est inspiré essentiellement par : The road to lysosome-related organelles: Insights from Hermansky-Pudlak syndrome and other rare diseases (2019).
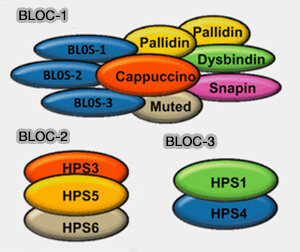
(Figure : vetopsy.fr d'après Flaumenhaft)
1. Les complexes BLOC (Biogenesis of Lysosome-related Organelles Complex) sont des complexes de sous-unités nécessaires à la biogenèse normale des organites spécialisés du système endosomal-lysosomal (LRO), comme les mélanosomes ou les granules denses des plaquettes…
2. On trouve trois complexes BLOC :
Complexe BLOC-1
Structure de BLOC-1
1. BLOC-1 contient huit sous-unités (Assembly and Architecture of Biogenesis of Lysosome-related Organelles Complex-1 (BLOC-1) 2012) :
- BLOS1, BLOS2, BLOS3 (Identification of Snapin and Three Novel Proteins (BLOS1, BLOS2, and BLOS3/Reduced Pigmentation) as Subunits of Biogenesis of Lysosome-related Organelles Complex-1 (BLOC-1) 2004),
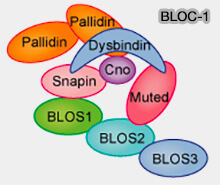
Complexes BLOC-1
(Figure : vetopsy.fr
d'après Mutagenetix)- muted (BLOC-1, a Novel Complex Containing the Pallidin and Muted Proteins Involved in the Biogenesis of Melanosomes and Platelet-dense Granules 2002),
- pallidine (Pallidin is a Component of a Multi-Protein Complex Involved in the Biogenesis of Lysosome-related Organelles 2002) ; il y aurait deux molécules de palladine (Platelet Secretion 2017),
- snapine (Snapin: a SNARE-associated protein implicated in synaptic transmission 1999),
- dysbindine (The dysbindin-containing complex (BLOC-1) in brain: developmental regulation, interaction with SNARE proteins and role in neurite outgrowth 2010).
2. Les noms de gènes humains pour chaque sous-unité sont référencés comme BLOC1S1-8 et leurs mutations provoquent des HPS (Hermansky-Pudlak syndrome).
BLOC-1 et tri des cargos
1. BLOC-1 est nécessaire pour fournir des cargos de protéines transmembranaires impliquées dans la synthèse de la mélanine, y compris TYRP1, OCA2, ATP7A et un petit sous-ensemble de TYR, des endosomes précoces aux mélanosomes via des transporteurs tubulaires (Voie 2).
2. BLOC-1 est exprimé de manière ubiquitaire et les rôles de BLOC-1 dans le trafic endosomal ont été identifiés dans d'autres types de cellules.
- Dans les cellules HeLa, BLOC-1 facilite le cycle de la transferrine à travers les endosomes de recyclage classiques.
- Dans les neurones, BLOC-1 et AP-3 collaborent pour assurer la médiation du cycle des cargos entre les endosomes et les vésicules synaptiques et peuvent réguler différemment le trafic des vésicules synaptiques dans différentes régions du cerveau (Roles of BLOC-1 and Adaptor Protein-3 Complexes in Cargo Sorting to Synaptic Vesicles 2009 et Hermansky-Pudlak Protein Complexes, AP-3 and BLOC-1, Differentially Regulate Presynaptic Composition in the Striatum and Hippocampus 2010).
Des souris déficientes en BLOC-1 avec une mutation inactivatrice de la dysbindine présentent des défauts dans la cinétique de libération des neurotransmetteurs (DTNBP1, a schizophrenia susceptibility gene, affects kinetics of transmitter release 2008.
- Plusieurs études ont suggéré que des variations dans les gènes codant pour la dysbindine et d'autres sous-unités de BLOC-1 sont corrélées à un risque accru de schizophrénie chez l'homme (The schizophrenia susceptibility factor dysbindin and its associated complex sort cargoes from cell bodies to the synapse 2011).
- Certains contestent cette conclusion (Dysbindin-containing complexes and their proposed functions in brain: from zero to (too) many in a decade 2011 avec une figure intéressante de ses partenaires potentiels).
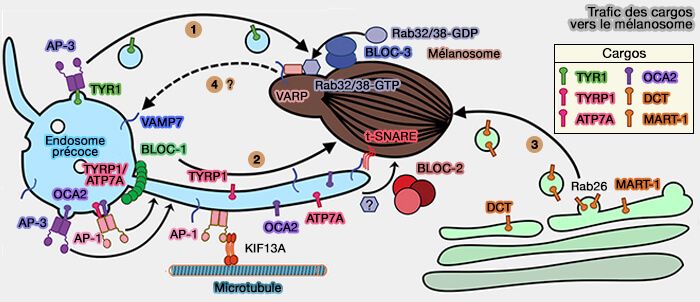
(Figure : vetopsy.fr d'après Bowman et coll)
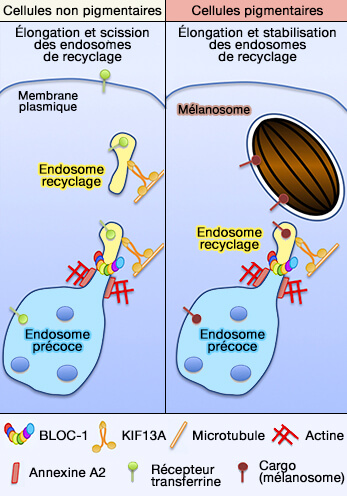
3. BLOC-1 se localise et est nécessaire à la formation des transporteurs tubulaires endosomaux qui fusionnent avec les mélanosomes pour livrer la cargaison.
Ces transporteurs ont des caractéristiques des endosomes de recyclage et régulent la formation de transporteurs tubulaires (AP-1 and KIF13A coordinate endosomal sorting and positioning during melanosome biogenesis 2009).
- BLOC-1, comme BLOC-2 dans les mélanocytes, se localise dans les endosomes tubulaires, et une cohorte de BLOC-1 et BLOC-2 interagit physiquement, suggérant que BLOC-1 et BLOC-2 fonctionnent dans la même voie (BLOC-1 Interacts with BLOC-2 and the AP-3 Complex to Facilitate Protein Trafficking on Endosomes 2006).
- Ce processus reflète une spécialisation de type cellulaire de la machinerie endosomale de recyclage, peut-être par le biais de la régulation par des composants spécifiques de type cellulaire encore indéfinis tels que les protéines Rab.
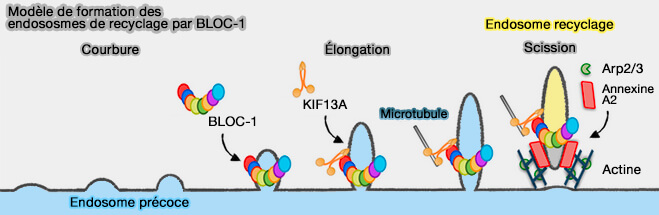
(Figure : vetopsy.fr d'après Delevoye et coll)
BLOC-1 et microsquelette
1. Les sous-unités de BLOC-1 forment une chaîne linéaire flexible, ce qui pourrait être cohérent avec les interactions de BLOC-1 avec des membranes tubulaires courbes (Assembly and Architecture of Biogenesis of Lysosome-related Organelles Complex-1 (BLOC-1) 2012 et Functional Characterization of Wiskott-Aldrich Syndrome Protein and Scar Homolog (WASH), a Bi-modular Nucleation-promoting Factor Able to Interact with Biogenesis of Lysosome-related Organelle Subunit 2 (BLOS2) and γ-Tubulin 2010).
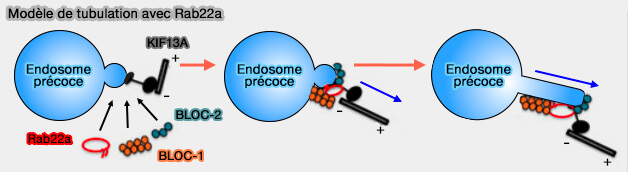
(Figure : vetopsy.fr d'après Shakya et coll)
Dans les mélanocytes et les cellules HeLa, BLOC-1 s'associe au moteur de la kinésine-3, KIF13A qui (AP-1 and KIF13A coordinate endosomal sorting and positioning during melanosome biogenesis 2009 et Recycling endosome tubule morphogenesis from sorting endosomes requires the kinesin motor KIF13A 2014) :
- relie peut-être les tubules membranaires au cytosquelette des microtubules, et favorise les réarrangements d'actine pour stabiliser et allonger les tubules (BLOC-1 Brings Together the Actin and Microtubule Cytoskeletons to Generate Recycling Endosomes 2016),
-
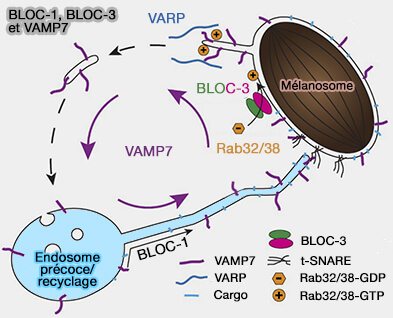
BLOC1, BLOC-3 et VAMP7
(Figure : vetopsy.fr d'après Delevoy et coll)
2. Ce complexe intervient dans la tubulation des endosomes précoces pour générer des endosomes de recyclage grâce à Rab22a ( Rôles de Rab22a dans le recyclage endocytaire).
Rôles de Rab22a dans le recyclage endocytaire).
Certaines de ces interactions peuvent être coordonnées par la petite GTPase Rab22 (Rab22A recruits BLOC-1 and BLOC-2 to promote the biogenesis of recycling endosomes 2018).
3 BLOC-1 favorise les réarrangements d'actine pour stabiliser et allonger les tubules (BLOC-1 Brings Together the Actin and Microtubule Cytoskeletons to Generate Recycling Endosomes 2016).
La perte de BLOC-1 :
- entraîne l'épuisement du complexe de nucléation de l'actine, Arp2/3.
- interagit avec le complexe WASH, un activateur Arp2/3, pour réguler le tri endosomal de la phosphatidylinositol-4-kinase de type IIɑ (PI4KIIɑ), qui possède un site de liaison avec AP-3, et qui produit du PI(4)P à partir du phosphatidylinositol (PtdIns) dans plusieurs types de cellules (Endosome-ER Contacts Control Actin Nucleation and Retromer Function through VAP-Dependent Regulation of PI4P 2016).
BLOC-1 possède un rôle dans la liaison du cargo au cytosquelette, actine et microtubules, avec des implications potentielles pour la fonction neuronale.
BLOC-1 et protéines SNARE
1. BLOC-1 pourrait également réguler la fusion membranaire médiée par les protéines SNARE.
- La pallidine interagit avec la sous-unité endosomale Qa-SNARE, la syntaxine 13 (The dysbindin-containing complex (BLOC-1) in brain: developmental regulation, interaction with SNARE proteins and role in neurite outgrowth 2010).
-
BLOC1, BLOC-3 et VAMP7
(Figure : vetopsy.fr d'après Dennis et coll)
2. VAMP7, la R-SNARE endo-lysosomale, requise pour la fusion des tubules dépendants de BLOC-1 avec des mélanosomes, se localise dans les vésicules synaptiques contenant AP-3 dans les cellules neuronales, suggérant un rôle plus large pour BLOC-1 dans le tri SNARE ( Vamp7 et BLOC).
Le tri de VAMP7 nécessite :
- soit la reconnaissance de VAMP7 par la sous-unité AP-3δ d'AP-3,
- soit celle de STX13 par la sous-unité pallidine de BLOC-1, mais pas les deux.
Les mélanocytes exprimant à la fois les variants AP-3δ et pallidine qui ne peuvent pas se lier à leurs protéines SNARE respectives sont hypopigmentés et ne parviennent pas à trier la cargaison dépendante de BLOC-1, STX13 ou VAMP7. Cependant, la liaison SNARE n'influence pas la fonction BLOC-1 dans la génération de supports de transport tubulaires (A BLOC-1–AP-3 super-complex sorts a cis-SNARE complex into endosome-derived tubular transport carriers 2021)
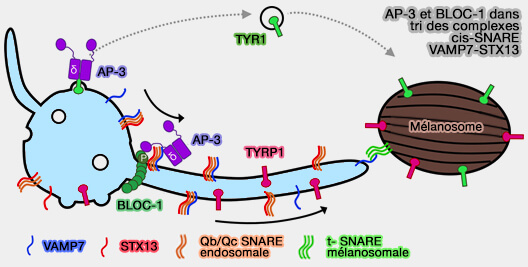
(Figure : vetopsy.fr d'après Bowman et coll)
Remarque : En plus de ces effets directs sur les protéines SNARE, la dysbindine de BLOC-1 peut se lier directement au NSF (N-éthylmaléimide Sensitive Fusion protein), l'AAA+ ATPase homohéxamerique impliquée dans le désassemblage du complexe SNARE pour réguler l'activité neuronale (The N-Ethylmaleimide-Sensitive Factor and Dysbindin Interact To Modulate Synaptic Plasticity 2015).
BLOC-1 versus BORC
1. Le complexe BORC (BLOC-One-Related Complex) contient trois sous-unités de BLOC-1, BLOS1, BLOS2 et la snapine.
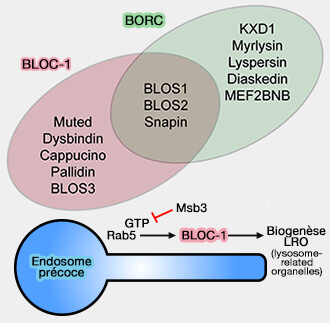
(Figure : vetopsy.fr d'après Langenmeyer et coll)
BORC contient également cinq sous-unités supplémentaires (BORC, a Multisubunit Complex that Regulates Lysosome Positioning 2015) :
- KXD1,
- myrlysine,
- lyspersine,
- diaskedine,
- MEF2BNB214.
Les gènes humains codant pour les cinq sous-unités spécifiques de BORC sont nommés BORCS4-8.
2. BORC est associé à la membrane des endosomes tardifs/lysosomes et recrute la petite GTPase Arl8b ( transport antérograde des endosomes tardifs/lysosomes) :
- pour remplacer Rab7,
- pour promouvoir le mouvement des lysosomes dépendant de la kinésine vers la périphérie cellulaire.
L'Arl8 actif, via l'adaptateur SKIP, recrute ensuite les moteurs de microtubules contenant la kinésine-1 contenant KIF5B ou la kinésine-3 contenant KIF1A ( recrutement des kinésines).
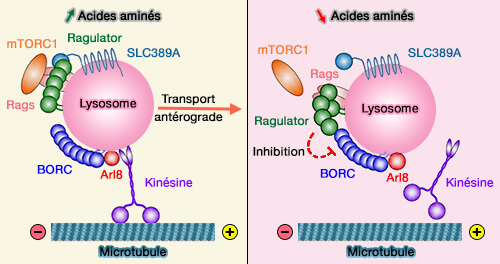
(Figure : vetopsy.fr d'après Pu et coll)
3. BORC interagit également avec Ragulator pour séquestrer les lysosomes dans la région périnucléaire en réponse à de faibles niveaux de nutriments dans les axones distaux ( positionnement des lysosomes régulé par les nutriments).
4. BORC, par l'activation d'Arl8, favorise l'acquisition du complexe HOPS, processus montré pour les autolysosomes (BORC coordinates encounter and fusion of lysosomes with autophagosomes 2017).
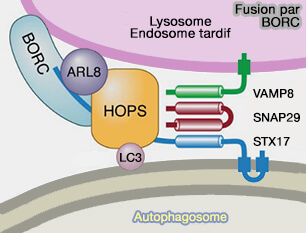
(Figure : vetopsy.fr d'après Pu et coll)
4. Le fait que BORC et BLOC-1 partagent trois sous-unités et que BORC n'ait été découvert que récemment a semé une certaine confusion dans la littérature concernant la fonction de chaque complexe.
a. Les inactivations de la snapine et de BLOS1 ont été attribuées en partie à BLOC-1, mais sont plus probablement dues au rôle de BORC dans la motricité et la fonction des lysosomes.
- L'inactivation du gène codant pour la snapine perturbe le transport rétrograde des vésicules synaptiques, des endosomes tardifs, et altère la fonction lysosomale dans les neurones de souris embryonnaires (Axonal autophagosomes recruit dynein for retrograde transport through fusion with late endosomes 2015).
- L'expression de mutants liant la dynéine à la snapine provoque le piégeage des vésicules synaptiques dans les terminaisons synaptiques (Regulation of synaptic activity by snapin-mediated endolysosomal transport and sorting 2016).
- L'épuisement de BLOS1 altère la dégradation lysosomale de EGFR, le récepteur du facteur de croissance épidermique (Biogenesis of Lysosome-related Organelles Complex-1 Subunit 1 (BLOS1) Interacts with Sorting Nexin 2 and the Endosomal Sorting Complex Required for Transport-I (ESCRT-I) Component TSG101 to Mediate the Sorting of Epidermal Growth Factor Receptor into Endosomal Compartments 2014).
b. La relation entre BORC et BLOC-1 et un complexe de levure appelé BLOC-1 est également source de confusion, et il n'est pas clair si ce complexe correspond au BLOC-1 de mammifère, au BORC ou à un complexe hybride (The BLOC-1 complex promotes endosomal maturation by recruiting the Rab5 GTPase-activating protein Msb3 2013 et Yeast Homologues of Three BLOC-1 Subunits Highlight KxDL Proteins As Conserved Interactors of BLOC-1 2010).
Complexes BLOC-2 et BLOC-3
Biologie cellulaire et moléculaireMembrane plasmiqueNoyauCytoplasmeMitochondriesSystème endomembranaireRéticulum endoplasmiqueAppareil de GolgiEndosomesLysosomesPeroxysomesProtéasomesCytosquelettemicrofilaments d'actineFilaments intermédiairesMicrotubulesMatrice extracellulaireReproduction cellulaireBiochimieTransport membranaireMoteurs moléculairesVoies de signalisation