


Système endo-lysosomal
Trafic des endosomes/lysosomes
Trafic antérograde (centrifuge) lié aux microtubules
- Vue d'ensemble du système endomembranaire
- Endosomes
- Lysosomes
- Trafic des endosomes/lysosomes
- Fusion et fission membranaire
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

La distribution spatio-temporelle des organites endo-lysosomaux dépend du transport entraîné par les protéines motrices des microtubules essentiellement, i.e. kinésines et dynéines, et celles des filaments d'actine, i.e. les myosines, aux sites de contact membranaire (MCS).
Les organites se déplacent dans tout le cytoplasme le long du cytosquelette, i.e. microtubules et filaments d'actine, et ces mouvements sont finement régulés ( vue d'ensemble du trafic des endosomes).
vue d'ensemble du trafic des endosomes).
1. Les protéines motrices de microtubules transportent les organites rapidement sur de grandes distances et de manière bidirectionnelle.
- La plupart des kinésines déplacent les organites de l'extrémité moins vers l'extrémité plus (transport antérograde ou centrifuge).
- La dynéine provoque le déplacement en sens inverse, de l'extrémité plus vers l'extrémité moins (transport rétrograde ou centripète).
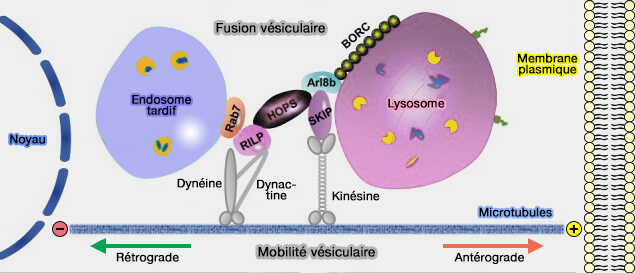
(Figure : vetopsy.fr d'après Khattern et coll)
2. Les moteurs de l'actine, i.e. les myosines, interviennent sur de courtes distances et de manière lente (transport lié au cytosquelette d'actine).
3. Le réticulum endoplasmique (RE), omniprésent dans tout le cytoplasme, joue un rôle majeur dans le contrôle du mouvement et du positionnement global des organites endo-lysosomaux ( trafic des endosomes lié au MCS du RE).
Généralités sur le transport antérograde (centrifuge)
Le transport antérograde (centrifuge), lié aux microtubules, utilise :
-
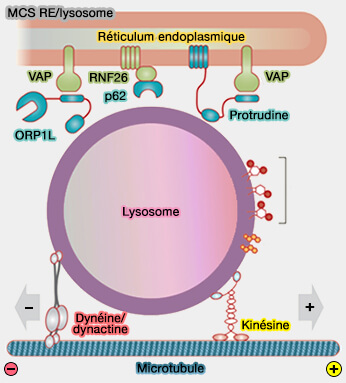
MCS RE/lysosome
(Figure : vetopsy.fr d'après Cabukusta et Neefjes) - les sites de contact membranaire (MCS) réticulum endoplasmique/endosomes.
Kinésines
Les mammifères codent pour environ 45 chaînes lourdes de kinésine (KIF) différentes, dont plusieurs entraînent le mouvement des organites endo-lysosomaux ( tableau).
En règle générale, une kinésine ne correspond pas à un organite particulier.

Les kinésines sont étudiées dans un chapitre spécial avec les moteurs moléculaires.
1. Dans certains cas, des organites distincts dépendent de la même kinésine pour leur mouvement.
La kinésine-3 (KIF16B) intervient dans le transport antérograde :
- des endosomes précoces (Modulation of Receptor Recycling and Degradation by the Endosomal Kinesin KIF16B 2005),
- des endosomes de signalisation SARA,
- des endosomes transcytosiques (The kinesin KIF16B mediates apical transcytosis of transferrin receptor in AP-1B-deficient epithelia 2013),
- des gouttelettes lipidiques (LD).
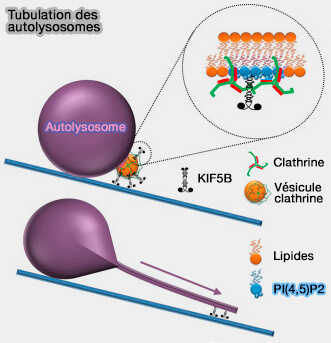
La kinésine-1 (KIF5B) est impliquée dans le transport antérograde :
-
Kinésines et trafic des lysosomes
(Figure : vetopsy.fr d'après Guardia et coll) - des mélanosomes (Rab1A regulates anterograde melanosome transport by recruiting kinesin-1 to melanosomes through interaction with SKIP 2015),
- des autolysosomes (reformation autophagique des lysosomes ou ALR).
La plupart des kinésines-1 dans les cellules animales sont tétramères, comprenant ( structure des kinésines) :
- deux sous-unités motrices identiques ou chaînes lourdes (KHC), i.e. ici KIF5, présentes en trois isoformes, i.e. KIF5A, B et C,
- deux chaînes légères (KLC) identiques, présentes en quatre isoformes, i.e. KLC1, 2, 3 et 4 (Axonal transport and neurological disease 2019).
2. Dans d'autres cas, les endosomes tardifs/lysosomes peuvent utiliser, outre KIF5B, plusieurs autres kinésines pour leur mouvement :
- la kinésine-2 - KIF3A - (Kinesin-2 is a Motor for Late Endosomes and Lysosomes 2005),
- la kinésine-3 - KIF1A et KIF1Bβ - (A Novel Kinesin-Like Protein, KIF1Bβ3 Is Involved in the Movement of Lysosomes to the Cell Periphery in Non-Neuronal Cells 2004),
- la kinésine-13 - KIF2A - (KIF2β, a new kinesin superfamily protein in non-neuronal cells, is associated with lysosomes and may be implicated in their centrifugal translocation 1998).
(Figure : vetopsy.fr d'après Du et coll)
KIF5B et KIF1A/KIF1Bβ couplent préférentiellement les endosomes tardifs/lysosomes à des pistes de microtubules distinctes.
Couplage des kinésines
Le couplage des organites endo-lysosomaux aux kinésines, i.e. adaptateurs d'organites, est médié par :
- les petites GTPases et leurs effecteurs,
- les phosphoinositides membranaires.
1. KIF16B, par exemple, est recruté dans les endosomes précoces par l'interaction de son domaine PX avec le PI(3)P, généré par VPS34, faisant partie des complexes phosphatidylinositol 3-kinase de classe III (PI3KC3), un effecteur de la petite GTPase Rab5.
2. Le recrutement de KIF5B et KIF1A/KIF1Bβ dans les endosomes tardifs/lysosomes, en revanche, dépend du complexe multi-sous-unité BORC et de la petite GTPase Arl8.
Transport antérograde (centrifuge) des endosomes tardifs/lysosomes
La kinésine-1 est recrutée, via les chaînes légères de la kinésine (KLC), par :
- Arl8 et SKIP/PLEKHM2 régulées par BORC,
-
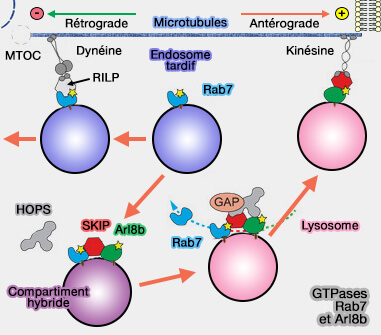
GTPases Rab7 et Arl8b
(Figure : vetopsy.fr d'après van der Kant et coll)
Arl8b
1. Arl8b (Arf-like GTPase8b ou ADP ribosylation factor like GTPase 8B) est une petite GTPase qui intervient dans la maturation des endosomes vers les lysosomes (Lysosomal Trafficking, Antigen Presentation, and Microbial Killing Are Controlled by the Arf-like GTPase Arl8b 2011).
2. Pour que Arl8b soit activée, elle doit :
- soit être recrutée dans des structures contenant Rab7,
- soit déjà se retrouver sur la même membrane en raison d'un événement de fusion antérieur (Arf-like GTPase Arl8: Moving from the periphery to the center of lysosomal biology 2015 et Emerging Roles of Arf-Like GTP-Binding Proteins: Fr om Membrane Trafficking to Cytoskeleton Dynamics and Beyond 2019).
(Figure : vetopsy.fr d'après Kendrick et coll)
Remarque : Arl8 peut aussi intervenir dans le trafic rétrograde ou centripète des endosomes tardifs/lysosomes par ses effecteurs qui favorisent le couplage des à la dynéine/dynactine en se liant à :
- PLEKHM1 (Pleckstrin Homology and RUN domain containing M1),
- RUFY3 et RUFY4 contenant les domaines RUN et FYVE en tant qu'effecteurs ARL8 pour le transport rétrograde le long des microtubules (RUFY3 and RUFY4 are ARL8 effectors that promote coupling of endolysosomes to dynein-dynactin 2022 et Bidirectional lysosome transport: a balancing act between ARL8 effectors 2022).
BORC
1. Le complexe BORC (BLOC-One-Related Complex) qui comprend 8 sous-unités, est associé à la membrane et recrute la petite GTPase Arl8b :
-
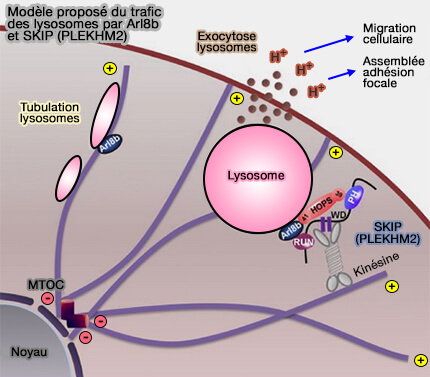
Modèle de fonctionnement des lysosomes par Arl8b
et SKIP (PLEKHM2)
(Figure : vetopsy.fr d'après Marwaha et coll) - pour promouvoir le mouvement des lysosomes dépendant de la kinésine vers la périphérie cellulaire.
BORC fonctionnerait comme un GEF pour Arl8b (BORC, a Multisubunit Complex that Regulates Lysosome Positioning 2015).
2. BORC partage un certain nombre de sous-unités avec BLOC1 (Biogenesis of Lysosome-related Organelles 1).
La structure de BORC et la comparaison BORC versus BLOC-1 sont étudiées dans un chapitre spécial.
3. L'activation d'Arl8b par BORC favorise l'acquisition du complexe HOPS, processus montré pour les autolysosomes (BORC coordinates encounter and fusion of lysosomes with autophagosomes 2017).
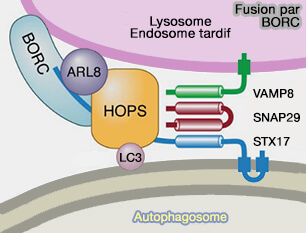
(Figure : vetopsy.fr d'après Pu et coll)
Arl8b pourrait être en concurrence avec RILP pour l'association avec le complexe HOPS (RILP interacts with HOPS complex via VPS41 subunit to regulate endocytic trafficking 2014).
SKIP/PLEKHM2
1. SKIP (Sif-A and Kinesin-Interacting Protein), un de ses effecteurs d'Arl8b, est aussi appelé PLEKHM2 ou Pleckstrin Homology and RUN domain containing M2 (The small GTPase Arl8b regulates assembly of the mammalian HOPS complex on lysosomes 2015).
SKIP, à ne pas confondre avec INPP5K, la phosphoinositide 5-phosphatases de type II, peut se lier à :
- Arl8b, via son domaine N-terminal RUN, i.e. domaine qui se lie aux petites GTPases, Arl8b se lie lui avec Vps41 du complexe HOPS,
- Rab7 via son domaine PH2 du domaine N-terminal, qui en contient 3. Ce domaine prendrait aussi en charge la liaison de SKIP avec Vps39 de HOPS.
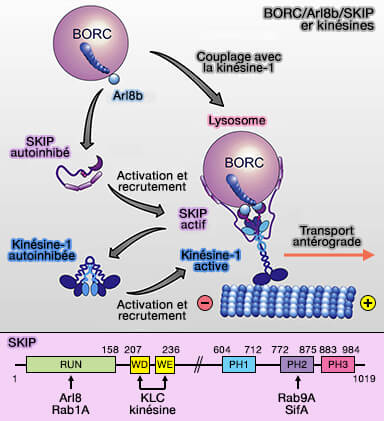
(Figure : vetopsy.fr d'après Keren-Kaplan et coll)
SKIP est auto-inhibé par l'interaction intramoléculaire de ses parties N- et C-terminales et serait activé par Arl8b (ARL8 Relieves SKIP Autoinhibition to Enable Coupling of Lysosomes to Kinesin-1 2021).
2. Le complexe Arl8b/SKIP/HOPS recrute le GAP TBC1D15 pour inactiver et éliminer Rab7 des membranes endosomales, donnant lieu à un compartiment spécifique à Arl8b ( Rab7 et TBC1D15).
3. SKIP est donc un effecteur négatif de Rab7 qu'il cible à différents emplacements cellulaires à des fins biologiques distinctes (SKIP-HOPS recruits TBC1D15 for a Rab7-to-Arl8b identity switch to control late endosome transport 2020).
Le mécanisme de commutation GTPase de Rab7 à Arl8b suit le paradigme de conversion de Rab5 à Rab7, définissant la prochaine étape de la maturation des organites endo-lysosomiaux.

SKIP régule de manière concomitante la liaison HOPS et le transport antérograde par la kinésine, indiquant que le moment de l'attache des vésicules est probablement couplé à leur transport sur les lysosomes.
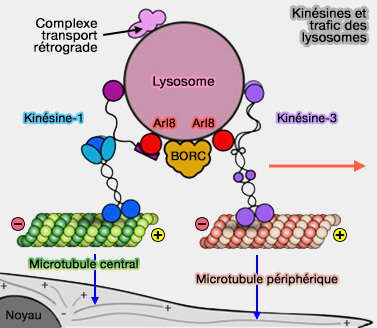
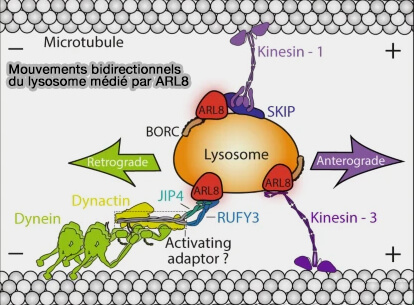
Recrutement des kinésines
Les interactions entre Arl8b et BORC favorisent le recrutement de (BORC Functions Upstream of Kinesins 1 and 3 to Coordinate Regional Movement of Lysosomes along Different Microtubule Tracks 2016) :
-
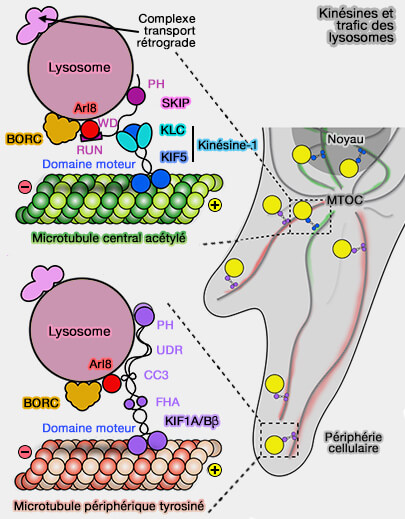
Kinésines et trafic des lysosomes
(Figure : vetopsy.fr d'après Guardia et coll) - la kinésine-3 dans les microtubules périphériques tyrosinés.
1. Le recrutement de la kinésine-1 (KIF5) implique des interactions :
- de Arl8 (les deux isoformes a ou b) avec le domaine RUN de SKIP,
- des domaines WD - tryptophane (W) et acide aspartique (D) - et WE - tryptophane (W) et acide glutamique (E) - dans une région non structurée de SKIP avec KLC2,
- de la chaîne légère de la kinésine (KLC2) avec KIF5B, la chaîne lourde de la kinésine, i.e. la kinésine-1 est un hétérotétramère KIF5B-KLC2.
2. Le recrutement de la kinésine-3, en revanche, est médié par une interaction directe entre Arl8 (a ou b) et le domaine CC3 (coiled-coil ou superhélice) des kinésine-3 KIF1Bβ ou KIF1A (The Balance between Capture and Dissociation of Presynaptic Proteins Controls the Spatial Distribution of Synapses 2013).
Remarques : la fonction des kinésines est contrecarrée :
- par le transport rétrograde (centripète) indépendant de BORC médié par la dynéine,
- par la protrudine qui se lie aux VAP ( inhibition du trafic des lysosomes par la protrudine).
Transport rétrograde (centripète) lié aux microtubules
Biologie cellulaire et moléculaireMembrane plasmiqueNoyauCytoplasmeMitochondriesSystème endomembranaireRéticulum endoplasmiqueAppareil de GolgiEndosomes/LysosomesPeroxysomesProtéasomesCytosquelettemicrofilaments d'actineFilaments intermédiairesMicrotubulesMatrice extracellulaireReproduction cellulaireBiochimieTransport membranaireMoteurs moléculairesVoies de signalisation