Les myosines diffèrent selon la vitesse de leur cycle ATPasique et la vélocité avec laquelle elles se déplacent le long du filament de myosine, mais aussi par :
le temps où la myosine reste attachée à l'actine (duty ratio),
l'efficacité avec laquelle l'actine peut déplacer l'ADP de la myosine (couplage thermodynamique et cinétique),
l'ampleur du mouvement de la myosine (sensibilité à la déformation).
Les classes de myosine peuvent contenir des moteurs de catégories différentes.
Moteurs rapides
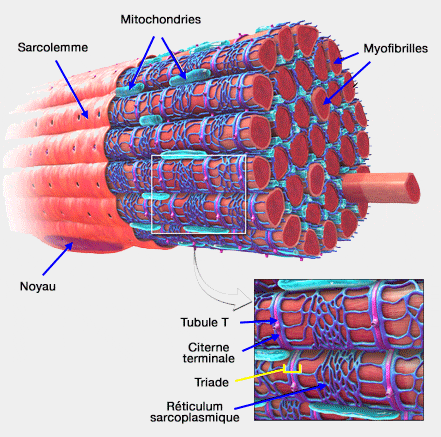
1. Ces moteurs rapides sont essentiels dans la contraction musculaire squelettique (liaison actine-myosine II) qui est décrite ici de manière très simplifiée.
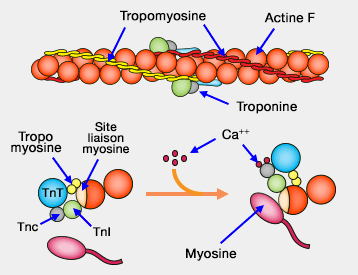
L'influx nerveux arrive dans la jonction neuromusculaire qui déclenche un potentiel d'action dans le sarcolemme (membrane plasmique) : ce message, transmis par les tubules transverses, ouvre les canaux calciques voltage-dépendants du réticulum endoplasmique qui libère donc des ions Ca++.
3. Dans les muscles lisses, les ions Ca++ libérés (qui doivent diffuser sur de longues distances car il n'y a pas de tubules transverses) se lient à la calmoduline.
L'activation de cet enzyme entraîne la phosphorylation de la RCL sur la Thr18 et la Ser19, ce qui a pour but de déplier la myosine pour former des filaments polaires et une possible contraction.
Ce type de régulation, diffusion lente du calcium et intervention de phosphorylation (une douzaine de facteurs comme MLCK, ROCK…), explique la lenteur du moteur et est dite régulation du filament épais.
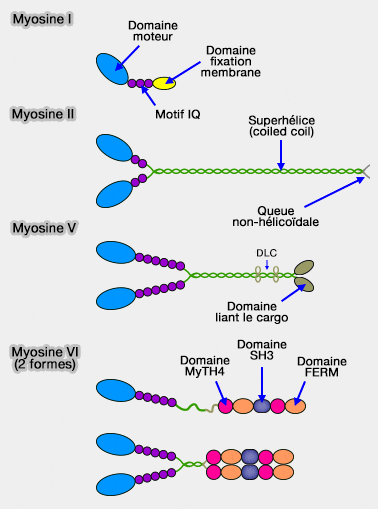
Structure de quelques myosines
(Figure : vetopsy.fr d'après Krendel)
Contrairement aux muscles squelettiques, la libération d'ions Ca++ est régulée par de nombreux signaux externes.
La noradrénaline, l'angiotensine, l'histamine… provoquent des changements de forme ou d'adhérence des cellules non-musculaires.
4. Plusieurs sites de phosphorylation sont présents près du domaine C-terminal des chaînes lourdes et jouent un rôle dans la dissociation des microfilaments d'actine ou dans la prévention de leur polymérisation chez la levure : leurs rôles doivent être précisés chez les mammifères.
Moteurs processifs
1. Les moteurs processifs, comme les myosines V, ne se détachent jamais de l'actine lors de son déplacement (elles se lient également aux microtubules).
En biologie moléculaire et en biochimie, la processivité est la capacité d'une enzyme à catalyser des réactions successives sur une même molécule, sans la relâcher.
2. Les myosines V comportent deux chaînes lourdes, un long cou constitué de 12 chaînes légères et une queue dimérisée pour transporter différents cargos (Myosin V from head to tail 2008).
Le temps de travail de 70% de ces myosines est lié à la libération très lente de l'ADP, ce qui implique que l'une des deux têtes de myosine est en permanence lié à l'actine.
a. Dans le modèle dit de la chenille, chaque tête se déplace de 36 nm pour rejoindre l'autre.
b. Dans le modèle main à main (animation ci-contre), l'une des têtes est attachée à l'actine et la deuxième se déplace de 72 nm, donc 36 nm devant l'autre, et ainsi de suite.
La myosine V reste donc toujours attachée du même côté du filament.
Cette propriété est essentielle pour le transport des organites, processus dans lequel lâcher le cargo serait dommageable. En effet, la myosine V est impliquée dans :
Leur rôle est important chez les levures bourgeonnantes ou les algues… où elles servent à la répartition des organites dans les cellules : la myosine V est aussi appelé myosine XI chez les plantes.
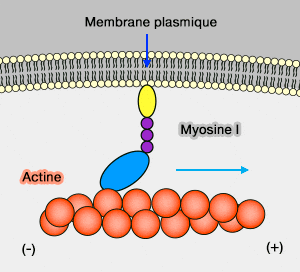
1. Les myosines I ont la particularité de se lier à l'actine comme les autres myosines par leur domaine moteur, mais aussi aux membranes.
Le génome des mammifères code huit isoformes de myosine-I (Myo1a-h) et les cellules immunitaires contiennent diverses combinaisons de myosine à queue courte comme Myo1c et Myo1g, et à longue queue comme Myo1e et Myo1f ( myosine I et phagocytose).
La queue des myosines I comprennent, en général, trois domaines :
TH1 participe à la liaison membranairecourte, chez la plupart (Myo1A, 1B, 1C, 1D, 1G, et 1H) qui possède un domaine PH (Pleckstrin Homology) qui se lie à ses motifs lipidiques (TH1 : Tail Homology domain 1),
TH2, riche en proline (PRD), contient un site de liaison à l'actine insensible à l'ATP,
TH3 est un domaine SH3 participe aux interactions protéine/protéine.
Les myosines I qui ne contiennent que le domaine TH1 sont appelées " à queue courte ", tandis que celles qui contiennent les trois domaines sont appelées " amiboïde " ou " à longue queue " (The myosin superfamily at a glance 2012
).
3. De nombreuses myosines I ont la particularité de maintenir la tension pendant de longues périodes sans l'hydrolyse de l'ATP.
Elles ont une affinité faible pour l'actine et donc restent peu de temps attachées et ne peuvent provoquer des contractions ou le transport de cargos sur de longues distances.
De ce fait, les myosines I régulent les interactions membrane-cytosquelette de manière dynamique pour pouvoir réagir aux variations des forces au cours de nombreux processus comme :
le maintien de la forme (tension) des membranes, comme dans le cortex cellulaire,



 cross-bridge dans la contraction musculaire).
cross-bridge dans la contraction musculaire).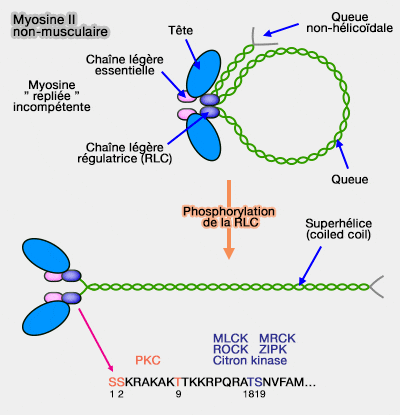

 myosine I et phagocytose).
myosine I et phagocytose).