Lors de la fusion membranaire, les protéines v-SNARE et les protéines t-SNARE, localisées sur des membranes séparées, forment un assemblage progressif en forme de fermeture éclair de quatre domaines SNARE, en général un de chaque sous-famille (Qa/Qb/Qc et R), formant le complexe trans-SNARE.
Le complexe SNARE est aussi appelé SNAREpin en anglais.
Lors de la fusion, le complexe trans-SNARE passe en position cis, i.e. complexe cis-SNARE dans lequel toutes les protéines SNARE sont positionnées sur la même membrane.
Vue d'ensemble du désassemblage
Par la suite, les complexes cis-SNARE doivent être désassemblés, afin de recycler les protéines SNARE pour un autre cycle de fusion membranaire, et ce par deux molécules :
la protéine α-SNAP/Sec17 (α-Soluble NSF Attachment Protein),
Les quatre molécules α-SNAP s'enroulent autour du complexe SNARE dans un arrangement de rotation symétrique, malgré le fait que le complexe SNARE lui-même n'est que pseudo-symétrique, i.e. les quatre hélices α sont constituées de différentes chaînes peptidiques.
L'anneau de rotation α-SNAP a une torsion vers la droite contrairement à la torsion vers la gauche du faisceau de quatre hélices α du complexe SNARE.
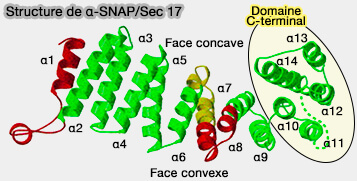
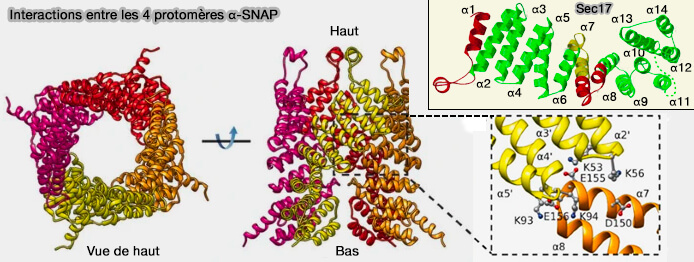
Structure et interactions des α-SNAP entre elles
(Figure : vetopsy.fr d'après Rice et coll et Zhou et coll)
Interactions entre les α-SNAP
1. L'interaction entre les α-SNAP se produit principalement au niveau de la partie N-terminale de la protéine, alors qu'il y a peu d'interaction au niveau de la #asnapstructureet implique principalement :
des résidus chargés négativement (D150, E155 et E156) des hélices α7 et α8 dans un protomère,
des résidus chargés positivement (K53, K56, K93 et K94) des boucles α2′-α3′ et α4′-α5′ dans le protomère adjacent.
2. Cette interaction électrostatique stabilise les quatre α-SNAP dans une structure serrée autour de la région C-terminale du complexe SNARE.
Cette interaction fait jouer aux α-SNAP :
un rôle dans le désassemblage du complexe SNARE.
un rôle possible dans l'assemblage α-SNAP-SNARE au stade précoce.
Remarque : l'absence d'interactions entre les α-SNAP au niveau de leur partie C-terminale peut expliquer la mobilité potentielle de ce domaine induite par l'hydrolyse de l'ATP médiée par la NSF.
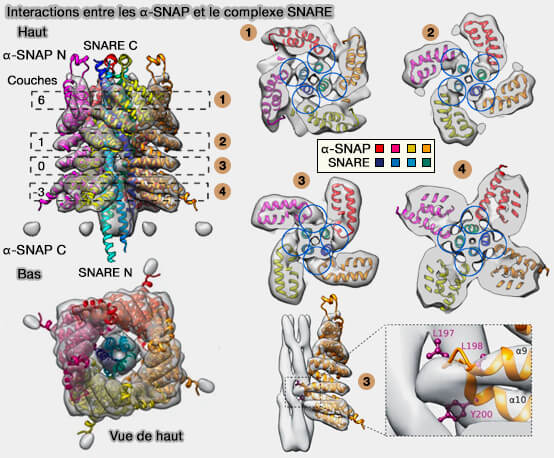
Interactions α-SNAP/SNARE
Interactions des α-SNAP avec le complexe SNARE
(Figure : vetopsy.fr d'après Zhou et coll)
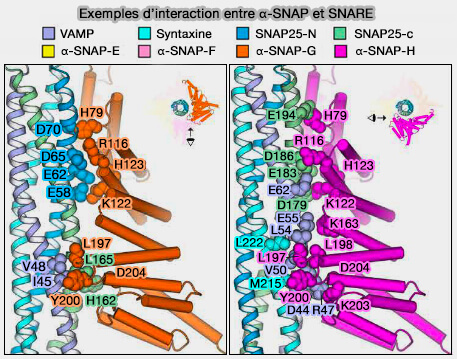
Chaque α-SNAP interagit avec trois des quatre chaînes du complexe SNARE, et ce par un ensemble identique de résidus.
Exemples d’interaction entre a-SNAP et SNARE
(Figure : vetopsy.fr d'après Huang et coll)
Par contre, différents résidus sont utilisés selon les SNARE individuelles.
a. Les deux sites d'interaction supérieurs, avec la couche 6 et 1, et le site d'interaction le plus bas (couche -3) se trouvent principalement sur la surface externe d'un brin hélicoïdal SNARE.
b. Le troisième site d'interaction, avec la couche 0, dans la partie inférieure forme la zone d'interaction la plus étendue avec les rainures du faisceau hélicoïdal SNARE.
Cette région d'interaction située à l'extrémité de l'épingle à cheveux en hélice α9/α10 des α-SNAP forme une structure en forme de cale se serrant entre les deux brins hélicoïdaux adjacents du faisceau à quatre hélices SNARE, qui semble correspondre aux couches -2/-3 du complexe SNARE.
Trois résidus hydrophobes hautement conservés (L197, L198 et Y200) de α-SNAP sont situés dans cette région, ce qui suggère qu'ils pourraient interagir avec le noyau hydrophobe du complexe SNARE et affaiblir l'interaction du faisceau à quatre hélices pour le désassemblage.
2. Les mutations des 12 résidus provoquent des perturbations dans le désassemblage du complexe SNARE.
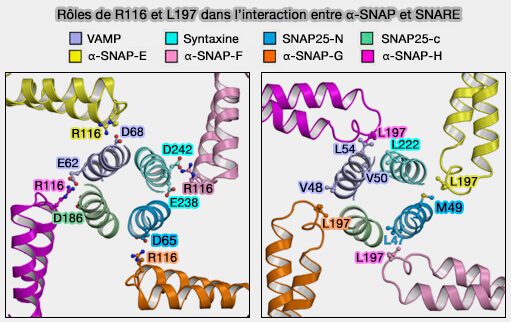
Rôles de R116 et L197 dans l'interaction entre α-SNAP et SNARE
(Figure : vetopsy.fr d'après Huang et coll)
Toutefois, R116 et L197 semblent jouer un rôle prépondérant, i.e. l'interaction entre les sites de la synaptobrévine /VAM, et les R116 et L197 de trois des quatre α-SNAP, E, G et H, en serait la cause.
Deux résidus R116 des α-SNAP-H et E interagissent avec deux sites de VAMP, E62 et D68, respectivement.
Deux résidus L197 de α-SNAP-G et H interagissent avec trois sites de VAMP, V48, V50 et L54, respectivement.




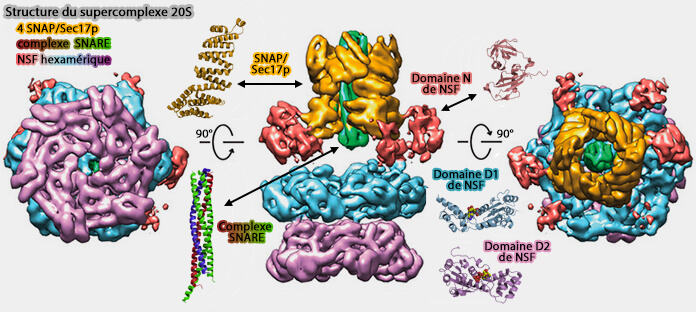

 Mechanistic insights into the SNARE complex disassembly (2019) pour plus de détails).
Mechanistic insights into the SNARE complex disassembly (2019) pour plus de détails).