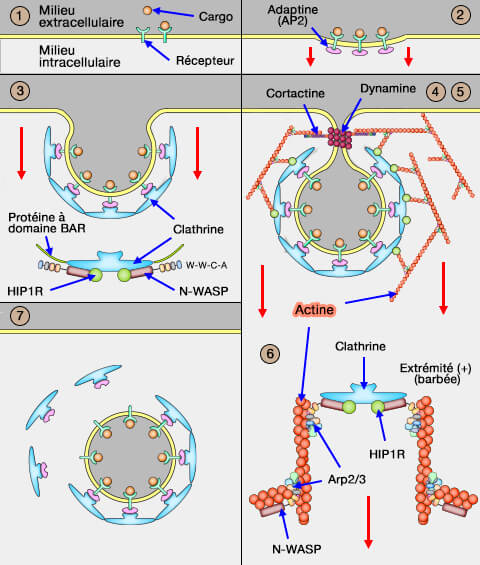
Dans l'endocytose clathrine-dépendante (CME), l'assemblage permet au puits formé par les adaptateurs de se recouvrir de clathrine (CCP ou Clathrin-Coated Pits).
3. L'assemblage permet au puits de se recouvrir de clathrine (CCP ou Clathrin-Coated Pits).
Un réseau de protéines (coat - " manteau, couche " ) à base de clathrine se forme à la face interne de la membrane plasmique pour former les CCS (Clathrin-Coated Structures), autre nom des CCP.
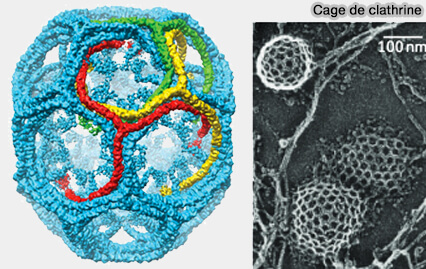
La formation des cages de clathrine (100 nm) et son renouvellement sont très rapides (2500 vésicules par minute avec une demi-vie d'une minute environ).
Clathrine vient du latin clastratus, grillage ou treillis.
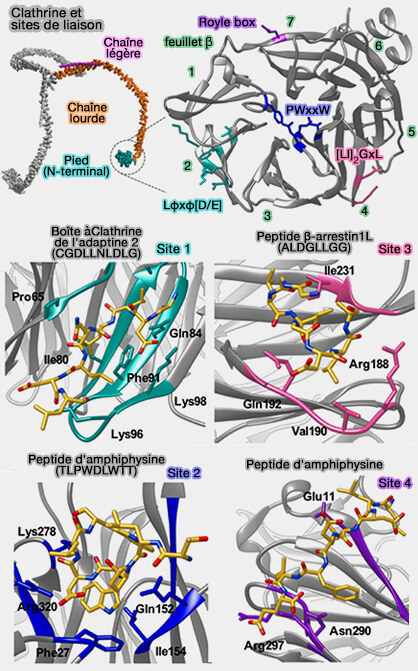
2. Le second site se lie aux protéines contenant la boîte W-motif (PWxxW) comme les amphiphysines 1 et 2.
3. Le troisième site est le deuxième site de liaison aux β-arrestines, en particulier longues comme la β-arrestine-1L, i.e. motif [L/I]2GxL, et un patch hydrophobe formé par les 4èmeet 5ème lames de clathrine.
Sites de liaison de la clathrine
(Figure : vetopsy.fr d'après Smith et coll)
4. Le quatrième site 4 est la Royle Box qui n'a pas encore de séquence d'interaction définie, mais qui entre en interaction avec la sous-unité β2 de l'AP-2.
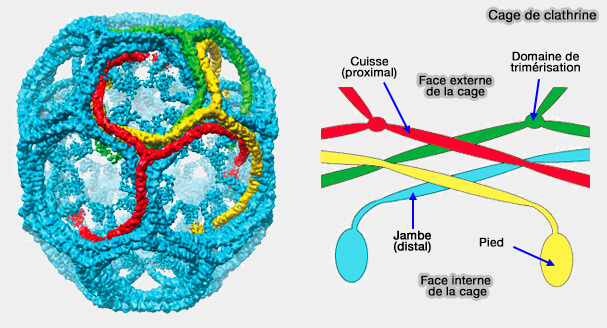
1. Suite au pied, on trouve la partie distale composée par :
la cheville (CHCR0, 1 et 2), qui interagit avec l'AP-2,
la jambe, partie distale (CHCR3, 4 et 5 N-terminal), un des sites de liaison du domaine J, par exemple de l'auxiline qui se lie aussi à Hsc70 pour stimuler l'activité ATPase.
3. Puis, la partie proximale est composée par :
le genou (CHCR5 C-terminal),
la cuisse, partie proximale (CHCR6 et 7), un des sites de liaison du domaine J.
4. Enfin, le domaine C-terminal est le siège de la trimérisation (entre les acides aminés 1488-1587), considéré comme un tripode hélicoïdal.
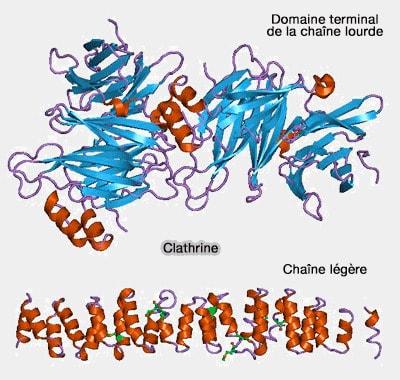
Chaîne légère (CLC)
Les vertébrés possèdent deux isoformes des chaines légères (CLC ou Clathrin Light Chain), i.e. CLCa et CLCb, codées par des gènes distincts, CLTA et CLTB.
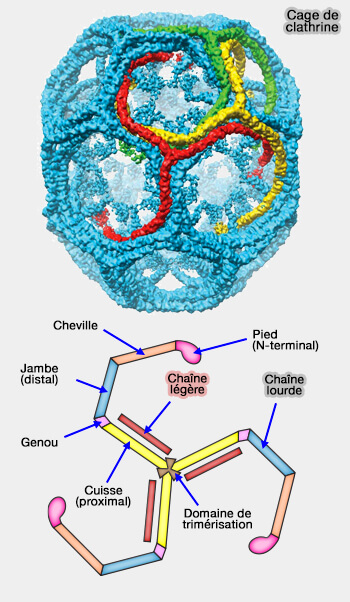
Les cages à clathrine, de 100 nm environ, lors de la formation de puits classiques dits aussi " canoniques " (Clathrin-Coated Pit ou CCP), sont construites par les interactions de nombreux triskélions (cf. figure du manteau de clathrine contenant 36 triskelia).
Cage de clathrine
(Figure : vetopsy.fr d'après N.V. Popova)
Pour fermer la vésicule, la cage doit incorporer 12 pentagones qui lui donnent sa courbure (et donc 6 hexagones).
La vésicule, en elle-même, de taille variable, en contient beaucoup plus, en formant un icosaèdre tronqué.
La formation de cages nécessite des domaines distaux correctement orientés par la liaison de leur domaine N-terminaux avec l'adaptateur.
Cage de clathrine
(Figure : vetopsy.fr d'après N.V. Popova)
3. Les pieds, domaines terminaux, sont dirigés vers l'intérieur, en direction du centre de la cage et se trouvent sous les sommets.
Ils prennent une forme de crochets qui permettent leur fixation à la couche interne formée par les protéines adaptatrices. Ils sembleraient interagir avec les chevilles des autres trimères.
En outre, leurs trois segments C-terminaux forment une cavité en forme d'entonnoir qui contient un motif QLMLT (résidus 1638–1642 chez les mammifères) qui permet une liaison optimale avec la protéine Hsc70, protéine de déshabillage des cages de clathrine.
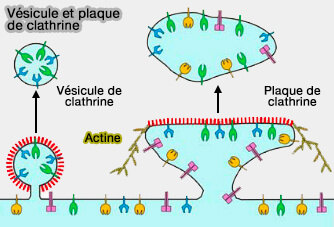
" Plaques " à clathrine
Les " plaques " non canoniques, formées surtout d'hexagones qui produisent un réseau de faible courbure ou même plat, ont encore un rôle ambigu.
Vésicule et plaque de clathrine
(Figure : vetopsy.fr d'après Traub)
Les adhérences focales digèrent l'ECM en créant des indices topographiques extracellulaires qui dictent l'emplacement futur des plaques recouvertes de clathrine.
Eps15 et Eps15R comme des régulateurs clés pour la formation de plaques recouvertes de clathrine sur les sites ECM localement remodelés.
Les plaques de clathrine agiraient comme des plateformes capables de recruter l'actine corticale ramifiée qui, à son tour, ancre les filaments intermédiaires à base de desmine, tous deux essentiels pour la formation et la fonction musculaire.
Remarque : les processus de formation des cages et des plaques sont distincts, i.e. il n'y a pas de transformation - montage, démontage - d'une à l'autre comme on le supposait, et leur relation, s'il y en a une, est pour l'instant obscure (Distinct Dynamics of Endocytic Clathrin-Coated Pits and Coated Plaques 2009).