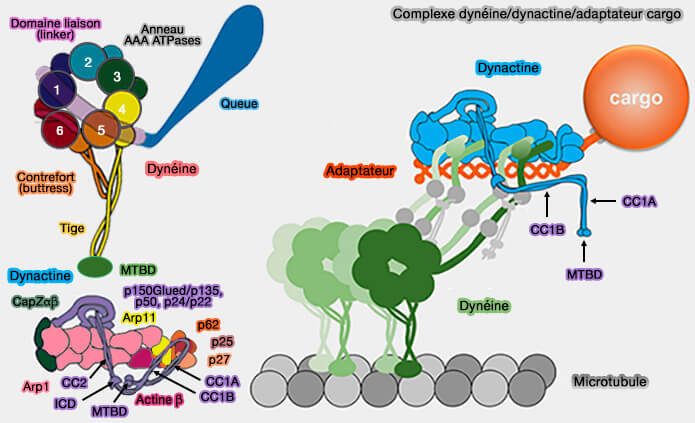
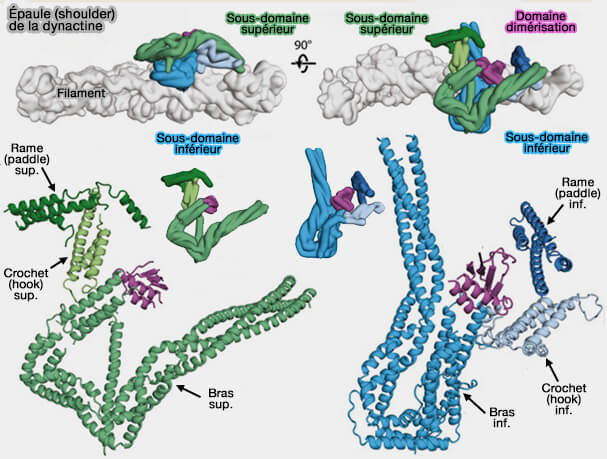
Ces protéines forment de longs faisceaux d'hélices alpha qui s'enroulent les unes sur les autres et entrent en contact avec le filament.
Les extrémités N-terminales des p50 émergent de l'épaule et recouvrent le filament, fournissant un mécanisme de contrôle de sa longueur.
Les extrémités C-terminales des p150glued, sont intégrées dans l'épaule, alors que les extrémités N-terminales sont projetés à l'extérieur et forment le bras.
2. Ces deux domaines, liés par un domaine de dimérisation, sont formés par deux bras contenant :
trois hélices α, i.e. hélices de p24 et p50-A/B,
un domaine hélicoïdal court, appelé crochet (hook), i.e. hélices de p50-A et d'une partie du domaine N-terminal de p150glued,
un autre domaine hélicoïdal court, appelé rame (paddle), i.e. hélices de p50-A/B,
Le domaine de dimérisation est formé par les quatre sous-unités.
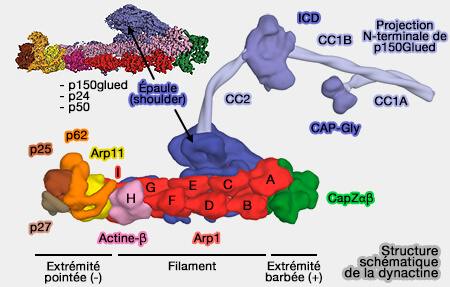
Épaule (shoulder) de la dynactine
(Figure : vetopsy.fr d'après Lau et coll)
Structure de l'épaule
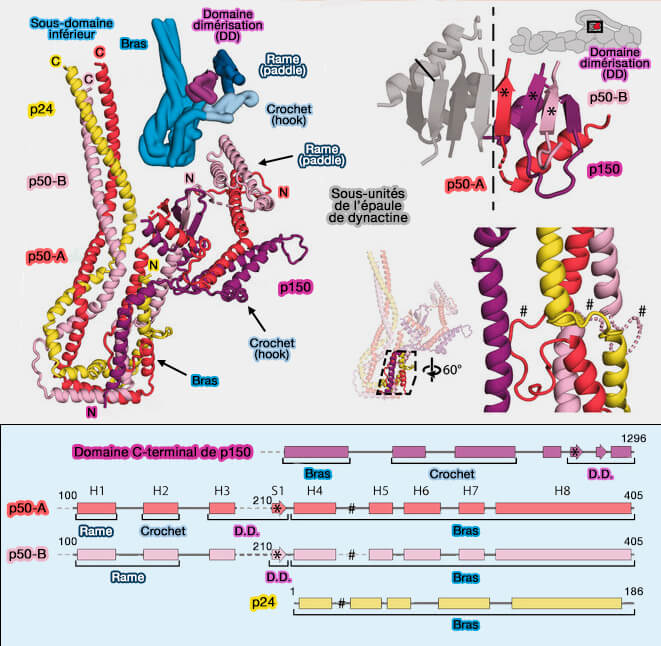
Partie C-terminale de p150
La partie C-terminale de p150 pénètre dans l'épaule autour du résidu 1096.
Elle se lie avec les parties de p50 et p24 au milieu du bras, ce qui suggère fortement qu'il s'agit d'une partie obligatoire de l'épaule de la dynactine.
Les résidus 1096–1140 courent le long du bras, établissant des contacts avec les sous-unités p50 et p24.
La section suivante de p150 forme une épingle à cheveux hélicoïdale qui représente les deux tiers du crochet.
La chaîne polypeptidique se replie alors de manière inattendue dans le domaine de dimérisation, contribuant à deux brins β et une hélice α (résidus 1253–1286).
p50
1. La région de p50-A/B qui est intégrée dans l'épaule comprend les résidus 100 à 405.
Les parties C-terminales de p50 forment un faisceau à trois hélices avec l'intégralité de p24 pour constituer le bras long.
Les deux p50 relient le bras au domaine de dimérisation..
2. Dans le crochet et la rame, les parties les plus N-terminales des deux p50 se replient dans différentes conformations, i.e. les résidus 1 à 100 dans chaque sous-domaine se projettent des côtés opposés des rames.
p50-B à l'extrémité N-terminale de l'autre côté du filament interagit avec les sous-unités Arp1-A, -C et -E (ER-1).
Les connexions du sous-domaine supérieur au filament sont faibles mais sur la base de la proximité, p50-A devrait se connecter aux sous-unités Arp-G et -I (ER-2), et p50-B à la sous-unité Arp-F (ER-4).
L'asymétrie dans l'épaule fait que chacune des quatre extrémités N-terminales est présentée de manière unique, afin d'interagir avec les huit sous-unités Arp1.
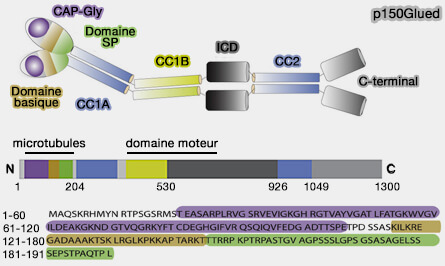
Bras : projection N-terminale de p150glued
Les derniers 1227 acides aminés des deux p150glued, composés d'environ 1300 résidus, qui se projettent en dehors de l'épaule (de 50 nm environ), sont constitués, chacun, par plusieurs domaines impliqués dans la liaison des microtubules.
2. Le deuxième site de liaison se situerait dans le domaine basique (S111-I116, A124-R132 et K144-T146).
Les domaines basique et riche en SP sont intrinsèquement désordonnés en solution et améliorent considérablement l'affinité de liaison aux microtubules.
p150glued conserve un degré élevé de flexibilité lors de la liaison aux microtubules, à l'exception de ces régions.





 Protéines de stabilisation : tropomyosine).
Protéines de stabilisation : tropomyosine).