


Petites GTPases
Effecteurs
Domaine GBD/PDB
/RBD
- En construction

Le domaine GBD (GTPase-Binding Domain), appelé aussi PBD (p21 binding domain) est un domaine de liaison domaine de liaison aux petites GTPases, en particulier de la famille Rho/Rac/Cdc42.
Les états inactifs du GDP et les états actifs liés au GTP sont reconnus par différentes protéines partenaires, permettant ainsi aux petites GTPases de fonctionner comme des commutateurs moléculaires.
L'état lié au GTP fournit une plate-forme pour l'interaction sélective avec les cibles en aval, les protéines dites effectrices.
Domaine RBD pareil que GBD ?
Les sous-unités alpha des protéines Ras et G hétérotrimériques sont des GTPases transductrices de signaux qui alternent entre des formes inactives liées au GDP et des formes actives liées au GTP. Les activités de ces GTPases sont régulées en partie par les protéines activatrices de la GTPase (GAP) qui stimulent l’hydrolyse des GTP, et les facteurs d’échange de nucléotides de guanine (GEF) qui stimulent la libération de GDP. Les GTPases Ras et G alpha sont des molécules de signalisation prolifiques qui interagissent avec un spectre de molécules effectrices et agissent par le biais de plus d’une voie de signalisation. Le domaine de liaison Ras (RBD) est un domaine indépendant d’environ 75 résidus, ce qui est suffisant pour la liaison dépendante du GTP de Ras et d’autres G alpha GTPases. Le domaine RBD peut être présent seul ou en tandem et il peut être associé à de nombreux autres domaines, tels que PDZ, RGS, PID, PH, C1, DH ou protéine kinase [1]. Structurellement, le domaine RBD de Raf-1 se compose d’une nappe bêta mixte à cinq brins avec une hélice alpha interrompue et deux petites hélices alpha supplémentaires. La structure du domaine RBD appartient au superfold du rouleau alpha/bêta de l’ubiquitine et est similaire à celle du domaine RA malgré l’absence d’identité de séquence significative. L’interaction majeure entre le domaine RBD de Ras et de Raf-1 se produit entre deux brins bêta antiparallèles : bêta 2 de Ras et bêta 2 de RBD [2]. Certaines protéines connues pour contenir un domaine RBD sont énumérées ci-dessous : - Vertébrique Raf proto-oncogène sérine/thréonine-protéine kinase. - Régulateur chez les mammifères de la signalisation des protéines G 12 (RGS12) et 14 (RGS14). - Invasion du lymphome T de mammifère et protéine induisant des métastases 1 (TIAM1). TIAM1 module l’activité des protéines de type Rho et relie les signaux extracellulaires aux activités cytosquelettiques. - Facteur d’échange de type Sif et Tiam1 de souris (STEF). - Drosophile LOCO-c1 et LOCO-c2, deux protéines nécessaires à la différenciation des cellules gliales. - Protéines de nature morte de drosophile (SIF) de type 1 et 2. Ils régulent la différenciation synaptique par l’organisation du cytosquelette d’actine, éventuellement en activant des GTPases de type Rho. Le profil que nous avons développé couvre l’ensemble du domaine RBD.
GBD (GTPase-binding domain)
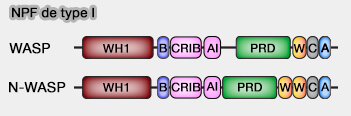
1. Le domaine de liaison aux petites GTPases, i.e. GBD (GTPase-Binding Domain), appelé aussi PBD (p21 binding domain) peut se diviser en deux domaines.
(Figure : vetopsy.fr)
a. Le domaine CRIB (Cdc42/Rac interactive binding motif) est la région N-terminale la plus conservée des domaines GBD/PBD.
- CRIB se compose d'environ 18 acides aminés avec la séquence consensus I-S-x-P-x-x-x-x-F-x-H-x-x-H-V-G.
- Bien que CRIB soit nécessaire pour la liaison à Cdc42 et à RAC, il n'est pas suffisant pour fournir une liaison à haute affinité.
b. Le domaine auto-inhibiteur (AI), appelé aussi IS (Inhibitor domain), moins bien conservé et localisé au niveau C-terminal de CRIB, est responsable du maintien des protéines dans un état basal, i.e. auto-inhibé.
Remarque : Le domaine GBD/PBD est souvent le synonyme de domaine CRIB.
2. Le domaine GBD est retrouvé dans de nombreux effecteurs des petites GTPases et se lie à Cdc42 et Rac sous leur forme GTP.
(Figure : vetposy.fr d'après Hoffmann et coll)
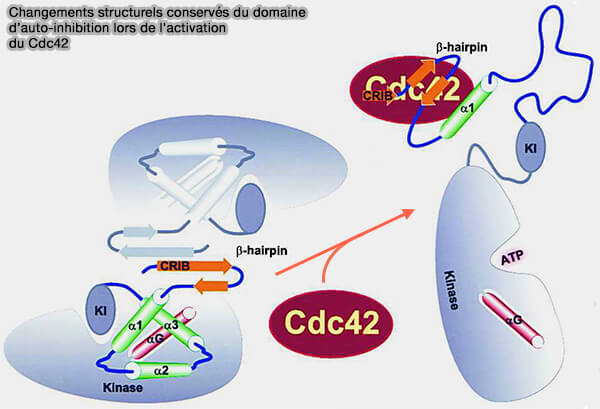
3. Le domaine inhibiteur est plié et composé de trois éléments architecturaux (Flipping the Switch Minireview The Structural Basis for Signaling through the CRIB Motif 2000).
a. Deux de ces éléments structurels sont dérivés du domaine AI/IS,
- une épingle à cheveux β N-terminale qui suit immédiatement le domaine CRIB conservé,
- un faisceau central de trois hélices α, i.e. α1, 2 et 3.
b. La troisième composante structurelle critique du pli inhibé est un segment hélicoïdal supplémentaire, αG, qui est distinct du domaine AI/IS.
- Dans le cas de WASP, cette hélice supplémentaire correspond au peptide limite de la région d'homologie de la cofiline, i.e. domaine C, qui conserve une activité de liaison vers le GBD/PBD.
- L'interaction du domaine AI/IS avec cette hélice est directement responsable de l'inhibition de l'activité de WASP (
 interactions de WASP avec Cdc42).
interactions de WASP avec Cdc42).
Vraisemblablement, l'interaction intramoléculaire du domaine SI avec l'hélice αG est incompatible avec la liaison au complexe ARP2/3 et, en l'absence de liaison de Cdc42, maintient WASP dans un état inactif.
Interactions avec les petites GTPases
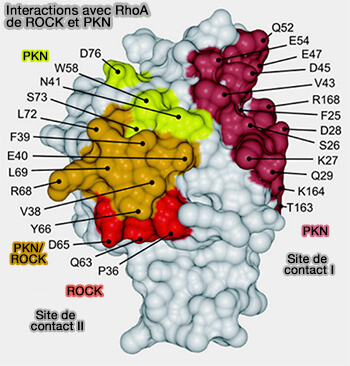
Les structures cristallines des domaines de liaison à la GTPase (GBD) de PKN et Rho kinase (ROCK) en complexe avec RhoA ont révélé que les domaines GDS forment une superhélice (coiled-coil) α-hélicoïdale.

Vous pouvez lire : The RHO Family GTPases: Mechanisms of Regulation and Signaling (2021).
(Figure : vetopsy.fr d'après Dvorsky et coll)
1. Pour ROCK1-GBD/RBD, i.e. Rho Bindig Domain (950-1012), une superhélice enroulée à gauche de 13 résidus dans sa partie C-terminale, i.e. 998LKTQAVNKLAEIM1010 est considérée comme la séquence minimale requise pour l'activité du motif d'interaction de Rho.
Elle se lie exclusivement aux régions de Switch et α2 de RhoA (Structural Insights into the Interaction of ROCKI with the Switch Regions of RhoA 2004).
2. En revanche, le complexe RhoA-PKN a deux sites de contact possibles sur RhoA (The Structural Basis of Rho Effector Recognition Revealed by the Crystal Structure of Human RhoA Complexed with the Effector Domain of PKN/PRK1 1999) et:
- Le site de contact 1 se compose des régions α1, β2/β3 et α5 de RhoA.
- Le site de contact 2 chevauche le site de liaison à ROCK.
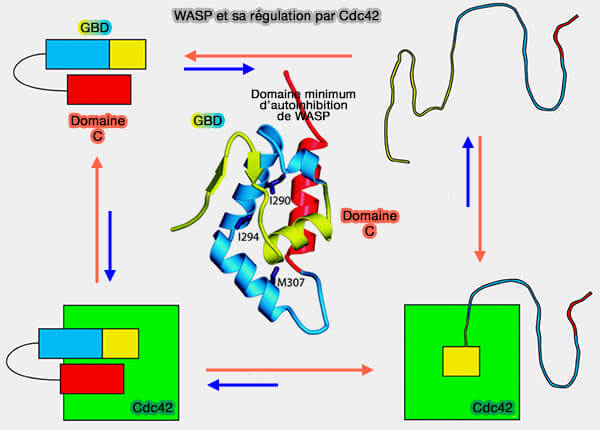
3. Les structures de Cdc42 en complexe avec des protéines effectrices contenant un motif de liaison Cdc42/RAC-interactive (CRIB) tel que PAK1 et WASP, montrent des contacts plus approfondis (The nucleotide switch in Cdc42 modulates coupling between the GTPase-binding and allosteric equilibria of Wiskott–Aldrich syndrome protein 2005).
a. Plus précisément, GBD :
- lie son épingle à cheveux β et à son hélice α-terminale aux régions α1, Switch I et II,
- s'enroule autour des régions α5 et β2 de la GTPase avec son terminal N étendu, qui englobe le domaine CRIB.
(Figure : vetposy.fr d'après Leung et coll)
b. Le domaine B de WASP, immédiatement en amont du domaine CRIB, génère des forces électrostatiques favorables à des résidus de glutamate uniques dans Cdc42 (Glu49, Glu171 et Glu178) qui contrôlent la réaction d'association WASP-Cdc42 accélérée (An Electrostatic Steering Mechanism of Cdc42 Recognition by Wiskott-Aldrich Syndrome Proteins 2005 et Functional Dysregulation of CDC42 Causes Diverse Developmental Phenotypes 2015).
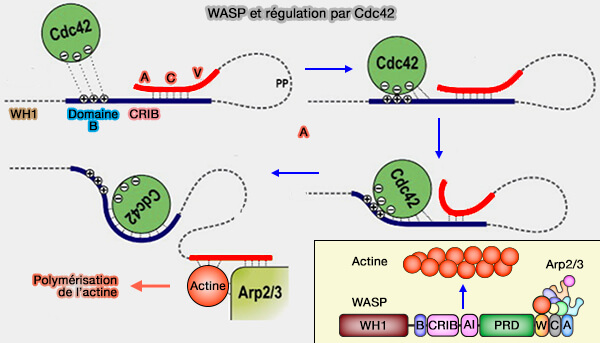
(Figure : vetpopsy.fr d'après Hemsath et coll)

Ce processus est une condition préalable à l'activation de WASP et une étape critique dans sa régulation temporelle et l'intégration de ses réponses cellulaires.

(Figure : vetpopsy.fr d'après Hemsath et coll)
Autres effecteurs
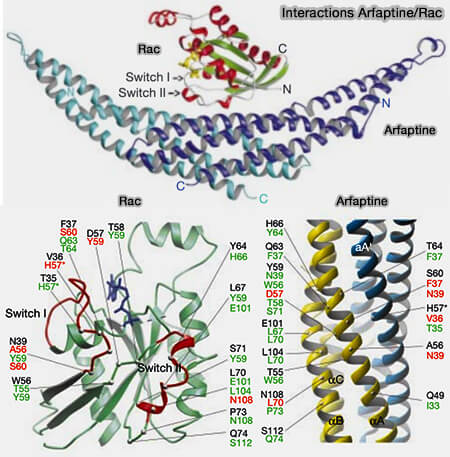
(Figure : vetopsy.fr d'après Tarricone et coll)
Deux autres effecteurs, l'arfaptine et P67phox, possèdent des structures différentes pour entrer en contact avec les GTPases.
Arfaptine
L'arfaptine forme un dimère en forme de croissant allongé formé par une superhélice (coiled-coil) de trois hélices α, formant un domaine BAR classique.
1. Elles entrent en contact avec les Switch I et II et α2 de Rac1, quel que soit son état lié aux nucléotides (The structural basis of Arfaptin-mediated cross-talk between Rac and Arf signalling pathways 2001).
Ces hélices imitent structurellement le domaine DH de TIAM1 (Structural mimicry of DH domains by Arfaptin suggests a model for the recognition of Rac–GDP by its guanine nucleotide exchange factors 2001).
2. Les arfaptines se lient à la petite GTPase Arf1 pour le bourgeonnement vésiculaire ( fonctions des domaineds BAR classiques).
2. L'arfaptine2 intervient dans la formation des autophagosomes ( translocation du complexe ULK).
P67phox
P67phox a un domaine hélicoïdal α qui se compose de quatre motifs de répétition tétratricopeptidique (TPR) de 34 résidus.
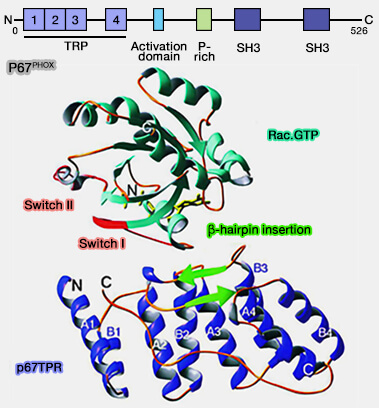
(Figure : vetopsy.fr d'après Lapouge et coll)
Ses motifs se lient à α1, aux résidus N-terminaux du Switch I, et aux boucles G3 et G5, mais pas là a région du Switch II ou les parties principales du Switch I (Structure of the TPR Domain of p67phox in Complex with Rac·GTP 2000).
Les régions de commutation pourraient être les sites de contact pour une troisième protéine associée au complexe Rac1-GTP-P67phox (Molecular basis for Rac2 regulation of phagocyte NADPH oxidase 2001).
Conclusion
Le mécanisme d'activation des structures complexes GTPase-effecteur est encore obscur.
Cependant, l'auto-inhibition intramoléculaire et l'exposition de leurs domaines fonctionnels sont nécessaires à leur fonctionnement.
Une caractéristique commune des complexes effecteurs est qu'à l'exception de P67phox, ils établissent tous un contact intensif avec les régions Switch/α2 des Rho GTPases, ce qui indique que cette région sert probablement de plate-forme pour la reconnaissance des effecteurs dépendants du GTP.
Retour aux petites GTPases de la famille Rho
En construction