Les fissions membranaires dans le trafic vésiculaire, i.e. voie sécrétoire et endocytose, doivent être à la fois spatialement et temporellement réglementées, afin que le cargo puisse être fidèlement livré au bon organite.
Les changements topologiques membranaires se produisent par des événements de fission qui font intervenir :
Si les mécanismes de fusion membranaire sont généralement bien compris, ceux de la fission sont plus difficiles à appréhender car ils sont dynamiques et transitoires.
En outre, de nombreux paramètres physico-chimiques interviennent dans la fission, comme la forme, la composition, la tension, la rigidité et la présence de protéines de la membrane…
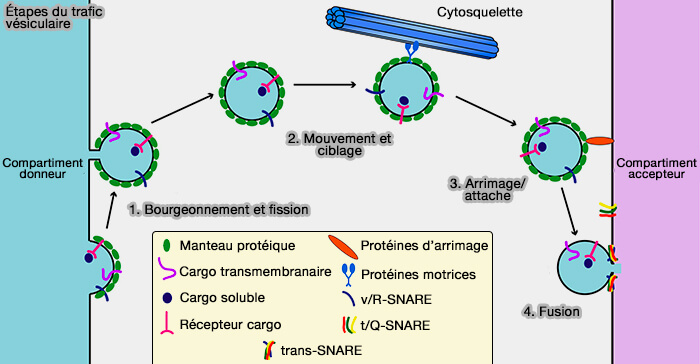
Étapes du trafic vésiculaire
(Figure : vetopsy.fr d'après Cai et coll)
1. Au sein de la cellule, le trafic vésiculaire contribue considérablement au transfert de protéines et de lipides entre les organites et entre les organites et la membrane plasmique.
Ces transporteurs (carriers) peuvent être ronds ou tubulaires et, quelle que soit leur morphologie, ils doivent contenir ou acquérir la machinerie nécessaire pour fusionner avec l'organite accepteur.
Les processus de fiction peuvent être divisés en quatre classes.
Vue d'ensemble
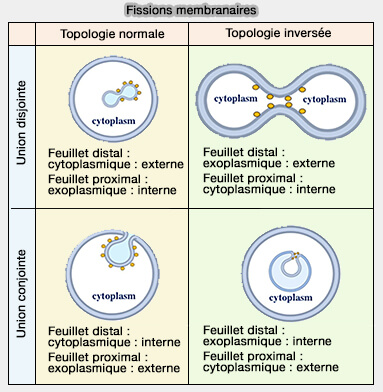
Lorsque la fission membranaire a lieu, la séparation des feuillets proximaux est suivie de la séparation des feuillets distaux.
Lors d'union disjointe compartiments membranaires, le feuillet externe est distal et le feuillet interne est proximal.
Lors d'union conjointe, le feuillet externe est proximal et le feuillet interne est distal.
1. La fission membranaire est dite à " topologie normale " lorsque les vésicules bourgeonnent de la membrane vers le cytosol.
a. Au cours de la fission topologique normale, le feuillet distal d'un compartiment membranaire est cytoplasmique, tandis que le feuillet proximal est exoplasmique.
Fissions membranaires
(Figure : vetopsy.fr d'après Zhukovsky et coll)
b. On assiste alors :
soit à une union disjointe de deux compartiments membranaires comme dans le cas de la formation des vésicules de l'appareil de Golgi, des exosomes ou des ectosomes,
soit à une union conjointe comme dans le cas de l'endocytose.







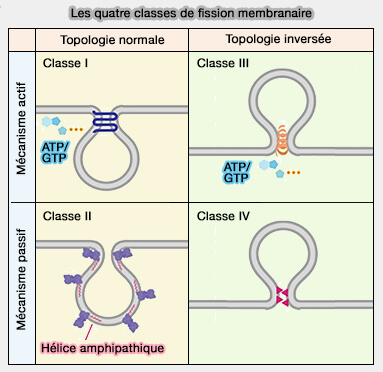
 protéines à AH provoquant la fission)…
protéines à AH provoquant la fission)…