


Motilité cellulaire
Migration cellulaire : vue d'ensemble
- En construction

La migration cellulaire, i.e. capacité des cellules à se mouvoir dans leur environnement, est un processus complexe effectué par tous les types de cellules nucléées à un moment donné de leur développement.
La migration cellulaire est donc un processus biologique fondamental.
- Le développement embryonnaire est caractérisé par des déplacements coordonnés cellulaires lors de la gastrulation à l'origine des trois feuillets primordiaux : les cellules, une fois en place, en général, ne se déplacent plus, sauf lors de réparation ou de tumeurs.
- La défense de l'organisme fait intervenir les mouvements des cellules immunitaires vers un foyer infectieux dans n'importe quel milieu.
- La cicatrisation effectuée par les cellules épithéliales induit des mouvements uniquement le long d'une membrane basale.
En outre, certaines cellules migrent très lentement comme les cellules endothéliales ou des cellules épithéliales intestinales.
- Les métastases tumorales relèvent aussi de la migration cellulaire.
Vue d'ensemble
La migration cellulaire est régulée.
1. D'une part, les mouvements doivent être :
a. dirigés vers un endroit précis et nécessitent une polarisation de la cellule, i.e. :
- un bord d'attaque (leading edge) en avant (dans le sens du mouvement),
- une partie arrière (queue).
b. coordonnés pour des déplacements collectifs.

Non seulement, cette migration peut faire intervenir des processus différents, mais elle doit être orchestrée en utilisant des voies de signalisation pour que l'ensemble des cellules se déplacent à un endroit précis et à un moment précis(Plasticity of cell migration: a multiscale tuning model 2009).
2. D'autre part, la migration cellulaire a été principalement étudiée in vitro sur un substrat fixe bi-dimensionnel.
a. In vivo, non seulement les cellules se déplacent (Life at the Leading Edge 2011) :
- à travers la matrice extracellulaire (MEC), mais aussi,
- les unes au-dessus des autres,
- les unes entre les autres,
- et même les unes au travers des autres.
b. Par exemple, les leucocytes traversent l'endothélium (diapédèse) :
- soit entre deux cellules endothéliales (voie paracellulaire), en dissociant une gap-jonction,
- soit en formant un canal à travers une cellule endothéliale (voie transcellulaire), en laissant la liaison intercellulaire intacte et en ouvrant un pore transmembranaire : on parle d'invadosome-like protrusions - ILP - (Mechanisms for transcellular diapedesis: probing and pathfinding by `invadosome-like protrusions' 2009).
3. La migration cellulaire implique généralement des changements drastiques de la forme des cellules.
La membrane plasmique forme différentes variétés de protrusions et, pour savoir de quoi on parle, certaines définitions sont indispensables.
Mécanismes généraux
1. Le mécanisme de migration cellulaire peut revêtir quatre formes principales (Life at the Leading Edge 2011).
a. Dans les filopodes et les lamellipodes, la polymérisation de l'actine pousse directement la membrane plasmique vers l'avant, i.e. :
- par des filaments parallèles dans les filipodes,
- par des filaments ramifiés dans la lamellipodes
b. Dans les invadopodes, la polymérisation de l'actine couplée avec des métalloprotéases dégradant la matrice extracellulaire (MEC) ouvre un chemin pour les cellules à travers la MEC.
c. Les blebs de membrane font avancer la membrane plasmique en utilisant une combinaison :
- de réseau contractile à base d'actomyosine,
- de détachement réversible de la membrane du cytosquelette d'actine cortical.
2. Chaque type de saillie nécessite la coordination d'un large éventail de molécules et les régulateurs de la dynamique du cytosquelette de signalisation.
En outre, ces différents processus sont susceptibles d'agir de concert pour déplacer les cellules dans des environnements complexes in vivo.
Quelques définitions indispensables
Les structures principales de la migration cellulaire sont stabilisées par des adhérences (Microtubules, compartiments cellulaires et GTPases Rho : ménage à trois pour une migration efficace 2001).
Structures protrusives adhérentes
(Figure : vetopsy.fr d'après Ridley)
Protrusions
Protrusions
classiques
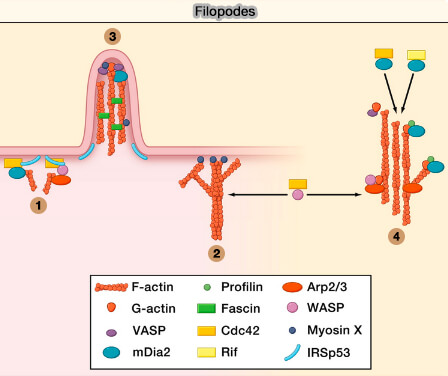
1. Les filopodes sont des protrusions ou extensions fines et cylindriques (en forme de doigt) de la membrane plasmique, contenant des filaments d’actine parallèles, qui explore l'environnement de la cellule et sont essentiels dans les cônes de croissance neuronale et l'angiogenèse des vaisseaux (Filopodia: The Fingers That Do the Walking 2007).
Les microspikes sont considérés comme des filopodes qui ne se projettent pas au-delà du bord de la cellule, contrairement à ces derniers : ils peuvent faire partie, comme les filopodes vrais, des lamellipodes ou alors être précurseurs des filopodes.
2. Les lamellipodes sont des protrusions larges, en forme d'éventail, correspondant à un réseau complexe de filaments d'actine ramifiés qui tirent les cellules à travers les tissus (Collective cell migration in morphogenesis, regeneration and cancer 2009)
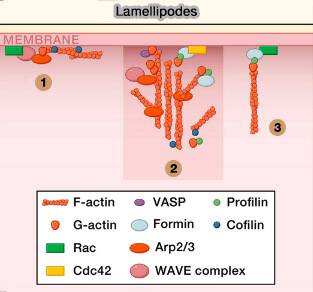
(Figure : vetopsy.fr d'après Ridley)
Les pseudopodes (ou " pseudo-pieds ") sont quant à eux des excroissances plus volumineuses que les lamellipodes, sous-tendues par un gel d’actine réticulée, que l’on trouve chez les amibes et les neutrophiles.
Les proportions de spectrine et d'actine varient selon le type cellulaire.
3. Les blebs, protrusions sphériques de la membrane sans cortex, dont la formation est liée à la pression intracellulaire, sont impliqués dans la direction de la migration pendant le développement.
Les cellules pourraient utiliser les deux modes de migration, les lamellipodes et les filopodes (mouvements mésenchymenteux) ainsi que les blebs (mouvements amiboïdes), selon l'environnement, i.e. (Control of Directed Cell Migration In Vivo by Membrane-to-Cortex Attachment 2010 et Cell mechanics control rapid transitions between blebs and lamellipodia during migration 2012) :
- la tension du cortex,
- la dynamique de l'actine,
- l'espace tridimensionnel ou pas.
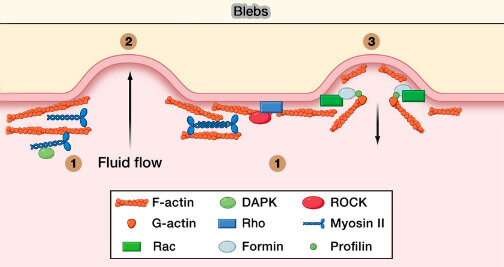
(Figure : vetopsy.fr d'après Ridley)
Protrusions pathologiques
1. Les invadopodes, sur lesquels nous n'insisterons pas dans ce site, sont impliqués dans les cancers invasifs avec métastases en détruisant focalement la matrice extracellulaire (MEC) par des enzymes adéquats (invadosomes at a glance, Matrix invasion by tumour cells: a focus on MT1-MMP trafficking to invadopodia et Actin machinery and mechanosensitivity in invadopodia, podosomes and focal adhesions dans Journal of Cell Science 2009).
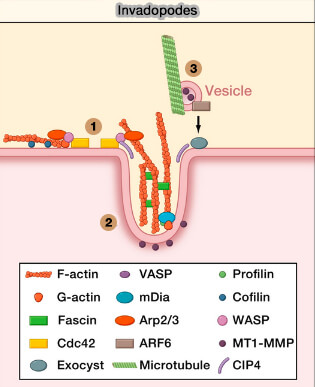
(Figure : vetopsy.fr d'après Ridley)
a. Leur formation faut intervenir des microtubules et des vésicules contenant (Life at the Leading Edge 2011) :
b. leur régulation est sous contrôle, pour la plus grande part, de la Rho-GTPase Cdc42 (Mechanisms for transcellular diapedesis: probing and pathfinding by `invadosome-like protrusions' 2009).
2. Les podosomes, typiques des macrophages et des ostéoclastes, sont des structures adhésives.
a. Ils sont formés d'un corps central de microfilaments d’actine entouré d’un anneau complexe qui comprend des intégrines et des protéines associées (paxilline, vinculine, taline…) se liant à l'actine et à la myosine.
b. Si de nombreux auteurs pensent que la structure des invadopodes et des podosomes sont identiques, d'autres pensent le contraire bien que les ressemblance soient nombreuses (WIP: WASP-interacting proteins at invadopodia and podosomes 2012).
Récemment, ces adhérences phagocytaires ont été comparées à des podosomes (Dynamic Podosome-Like Structures in Nascent Phagosomes Are Coordinated by Phosphoinositides 2019), des structures adhésives spécialisées à base d'actine capables de dégrader la matrice extracellulaire (Podosomes at a glance 2005).
À l'avenir, il sera intéressant de déterminer s'il existe des parallèles mécanistiques entre le rôle des myosines dans la régulation de la dynamique des podosomes (Tail domains of myosin-1e regulate phosphatidylinositol signaling and F-actin polymerization at the ventral layer of podosomes 2018) et leur contribution à la dynamique de l'actine au sein de la cupule phagocytaire.
Adhérences cellule/MEC
On en dénombre plusieurs catégories, mais pour notre sujet, ce sont surtout les points focaux qui nous intéressent ( autres adhérences cellule/MEC).
autres adhérences cellule/MEC).
1. Les contacts focaux (points focaux, adhérences focales ou plaques d’adhérence) sont des jonctions adhérentes ponctuelles entre la matrice extracellulaire (MEC) et la membrane plasmique de la cellule sous-jacente.
Ils réalisent le chaînon intermédiaire entre les molécules de la MEC et les microfilaments d’actine du cytosquelette, et en particulier sur les fibres de stress (tension).
2. Ces adhérences font intervenir essentiellement les CMAC (Cell–Matrix Adhesion Complexes) qui sont des complexes qui comprennent (Cell-matrix adhesion complexes: master control machinery of cell migration 2008) :
- des intégrines, en particulier l’intégrine alpha5-béta1,
- de nombreux adaptateurs intra-cytoplasmiques qui assurent le lien entre le domaine cytoplasmique des intégrines et les microfilaments d’actine,
- des protéines de signalisation.
Structures protrusives non adhérentes
Les replis membranaires (membrane ruffles), feuillets membranaires résultant de remaniements du cytosquelette d’actine sous-jacent à la membrane plasmique, se forment à la surface de la cellule et ne nécessitent pas de contacts focaux.
On en trouve deux sortes (Functions and Regulation of Circular Dorsal Ruffles 2012) :
- Les replis membranaires périphériques (PR : peripheral ruffles) se trouvent au bord d'attaque de la cellule en migration et qui croissent vers l'arrière en général.
- les replis membranaires dorsaux circulaires (CDR : Circular Dorsal Ruffle) se situent sur la face apicale de la cellule pour former, comme leur nom l'indique des structures circulaires pour inclure par exemple des récepteurs intramembranaires ou pour jouer un rôle dans la macropinocytose, dans la motilité et la migration cellulaires.
Vue d'ensemble de la migration cellulaire
Dans ce chapitre, nous étudierons la migration cellulaire par mouvement mésenchymental. Les mouvements par blebs sont envisagés dans le cortex cellulaire.
1. La migration cellulaire, contrôlée par de nombreuses molécules et voies de signalisation, est dépendante de l'actine (et des ABP - Actin Binding Protein -) qui peut se polymériser en différents réseaux.
- Le cortex cellulaire réticulé qui borde la membrane plasmique joue un rôle essentiel dans le maintien de la forme et des modifications cellulaires.
- Le reste de la cellule contient un réseau de filaments réticulés contenant des faisceaux contractiles (myosines), les fibres de stress.
2. Ce processus est régulé très rapidement, en particulier par les GTPases Rho, les phospholipides membranaires et les cycles de phosphorylation (kinases) et déphosphorylations (phosphatases), par :
- la polymérisation de l'actine grâce aux protéines de nucléation (complexe Arp2/3, formine),
- la dépolymérisation des microfilaments grâce à des protéines de fragmentation (ADF/cofiline en particulier), mais aussi grâce à la myosine (désassemblage des réseaux d'actine par la myosine).
Les propriétés des toxines dépolarisant l'actine (comme la latrunculine A) ont été utilisées pour étudier la vitesse de renouvellement de certains assemblages d'actine : elle est de 30 à 180 secondes dans le bord d'attaque des cellules mobiles et de 5 à 10 minutes dans les fibres de stress.
3. Cette migration cellulaire se fait en 4 temps :
- l'extension grâce des lamellipodes ou des filopodes,
- l'adhérence (adhésion) dans laquelle la protrusion adhère au substrat (points focaux),
- la translocation au cours de laquelle tout le contenu cellulaire se déplace vers l'avant,
- le détachement de la queue qui consiste à la séparation de la partie arrière de la cellule du substrat.
Extension
En construction