Les mécanismes de fission passive résultent de la réorganisation spontanée des lipides et/ou des protéines qui conduisent à la fission sans utilisation directe d'énergie.
Wedging versus Scaffolding mecanisms
(Figure : vetopsy.fr d'après Boucrot et coll)
Domaines lipidiques et fission
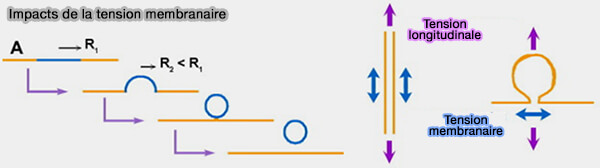
1. Les membranes peuvent comporter des domaines lipidiques différenciés par leur composition, conduisant à la coexistence de phases ayant des propriétés physiques différentes, i.e. épaisseur ou rigidité, qui provoquent des tensions de ligne ( courbure membranaire et lipides).
À leurs frontières, des perturbations énergétiquement défavorables apparaissent, provoquant une tension de ligne.
Le système membranaire tente de réduire ces perturbations en réduisant la longueur totale des frontières, i.e. l'énergie de ligne du système.
Si ce phénomène se produit dans une membrane tubulaire, i.e. col d'une vésicule invaginée, le diamètre du col est réduit par compression spontanée et peut conduire à la fission (bourgeonnement et fission des vésicules bi-phasiques 2005).
Forces en présence dans les phases de différentes compositions lipidiques
(Figure : vetopsy.fr d'après Lenz et coll)
2. Dans les membranes cellulaires in vivo, par la présence d'une grande variété de lipides, de protéines périphériques ou transmembranaires, la tension de la ligne est faible et n'est souvent pas suffisante pour conduire à la fission membranaire.
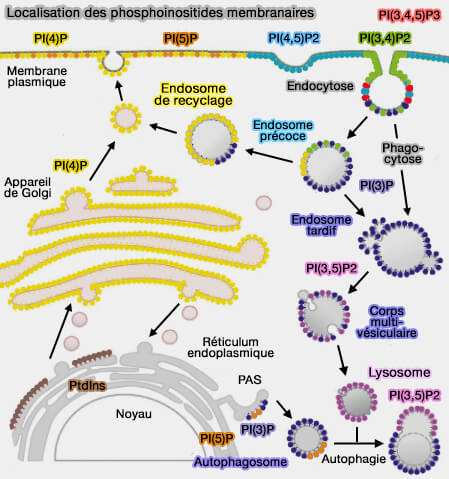
Localisation des phosphoinositides membranaires
(Figure : vetopsy.fr d'après Cernikova et coll)
a. La formation des domaines lipidiques membranaires est déclenchée par des signaux, qui conduisent, par exemple, à la phosphorylation ou à la déphosphorylation des phosphoinositides.
b. La tension de ligne est aussi contrôlée par des machineries actives, i.e. dépendante de l'énergie, comme le cytosquelette ( courbure membranaire et cytosquelette).
c. Dans certains cas, la fission passive pourrait être alimentée indirectement, via l'énergie utilisée dans la synthèse de ces cofacteurs lipidiques (Membrane scission driven by the PROPPIN Atg18 2017).
Crowding et fission
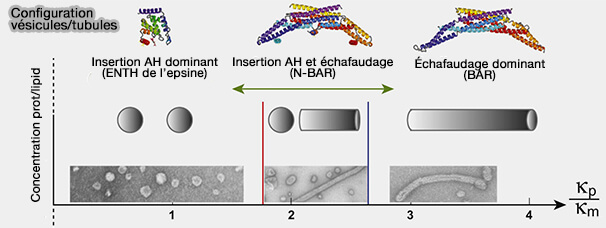
La courbure de la membrane est contrôlée par l'assemblage de nombreuses protéines avec des caractéristiques structurelles bien définies, notamment des revêtements sphériques en forme de réseau, des échafaudages tubulaires à domaines BAR et ESCRT, des hélices amphipathiques (AH) en forme de coin et des faisceaux, des filaments cytosquelettiques…
Comment l'environnement hétérogène et très encombré stériquement de la surface de la membrane cellulaire modifie-t-il la fonction des assemblages de protéines (Entropic Tension in Crowded Membranes 2012) ?
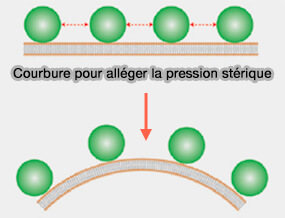
Courbure et crowding
(Figure : vetopsy.fr)
La pression latérale générée par les collisions entre les protéines liées à la membrane peut entraîner une courbure spontanée de la membrane et une tubulation.
Ce phénomène est indépendant de la manière dont les protéines sont attachées à la membrane, que ce soit via l'insertion de l'hélice amphipathique ou la liaison aux groupes de tête des phospholipides.
3. Dès lors, les protéines liées à la membrane plus grosses auront un impact plus important sur la pression stérique locale que les protéines liées à la membrane plus petites.
De nombreuses protéines impliquées dans le trafic membranaire contiennent des domaines volumineux intrinsèquement désordonnés, qui occupent des volumes considérablement plus importants par rapport aux motifs structurés de poids moléculaire équivalent (interactions dites " floues ", i.e. fuzzy en anglais).
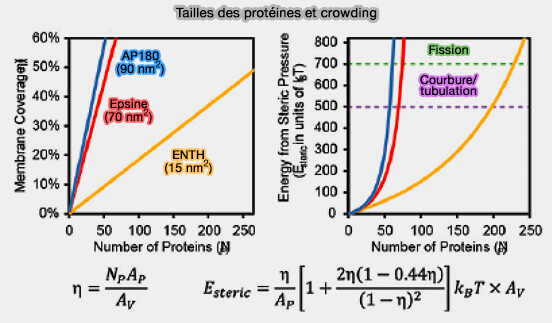
AP180 et l'epsine, qui contiennent chacun de grands domaines intrinsèquement désordonnés, sont beaucoup plus efficaces pour encombrer la surface de la membrane et entraîner le remodelage de la membrane par rapport au petit domaine ENTH globulaire (cf. figure ci-dessous).
Taille des protéines et crowding
(Figure : vetopsy.fr d'après Zeno et Stachowiak)
4. De même, la face luminale d'une structure membranaire incurvée, qui doit prendre une morphologie concave lors de la vésiculation membranaire, est généralement très encombrée par des domaines protéiques liés à la membrane tels que les ectodomaines des cargos transmembranaires, entre autres constituants.
La capacité du domaine ENTH à former des tubules membranaires et la vésiculation de la membrane pour générer des vésicules plus petites seraient corrélées à la couverture de la surface membranaire par les protéines plutôt qu'à l'insertion d'hélices amphipathiques dans les membranes.
Remarque : même une concentration élevée de GFP (Green fluorescent protein) est capable de transformer les surfaces membranaires planes en tubules à forte courbure.
L'équilibre de la pression stérique entre les surfaces membranaires opposées influence fortement la forme de la membrane et la fission.





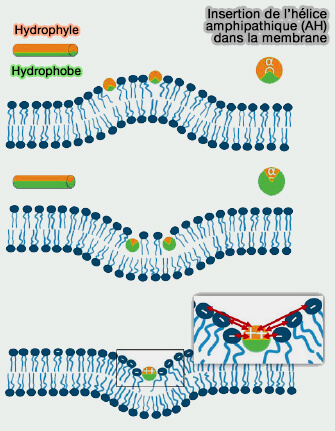
 protéines à AH provoquant la fission).
protéines à AH provoquant la fission).