


Microfilaments d'actine : polymérisation in vivo par le complexe Arp2/3
NPF : structure et localisation
- En construction

Les NPF (Nucleation Promoting Factor), classés en deux catégories selon la manière dont il interagissent et activent le complexe Arp2/3, diffèrent dans leur structure.
-
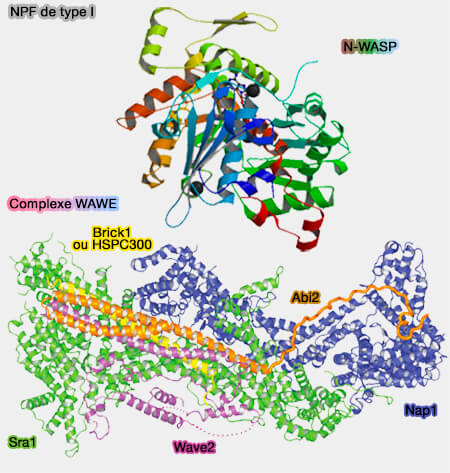
NPF de type I : N-WASP et le complexe WAVE
(Figure : vetopsy.fr) - Les NPF de type II (cortactine ou son homologue Abp1), ne possèdent pas de domaine VCA (
 protéines de stabilisation).
protéines de stabilisation).
Structure des NPF
Domaine C-terminal : VC/WCA
Cette famille comporte de nombreux membres qui possèdent tous un domaine N-terminal spécifique et un domaine C-terminal VCA/WCA (WH2 Connecteur Acid), indispensable à l'activation de l'Arp2/3.
Ces protéines intrinsèquement désordonnées (intrinsically disordered proteins) réglementent l'assemblage de l'actine de façon adaptative (WH2 domain : a small, versatile adapter for actin monomers 2002).
Le domaine VCA comprend trois domaines essentiels.
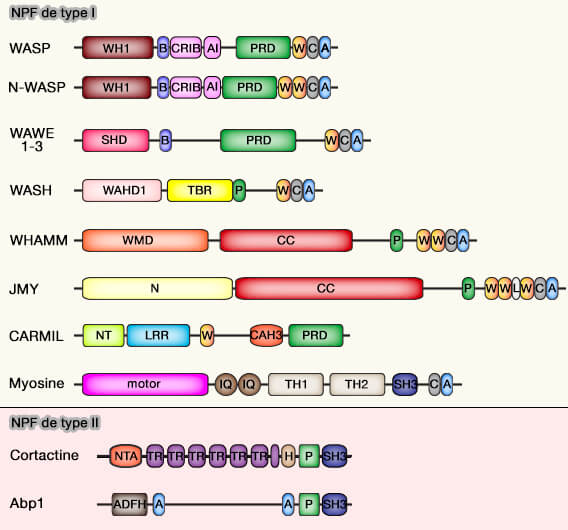
(Figure : vetopsy.fr)
Domaine WH2

Vous pouvez lire : The WH2 Domain and Actin Nucleation – Necessary but Insufficient (2016).
Vue d'ensemble
Le domaine WH2 (WASP-Homology 2) se lie à l'actine : il est aussi appelé V, pour verprolin homology, d'où le nom VCA (The β-Thymosin/WH2 Fold Multifunctionality and Structure 2007 et Structure, Function, and Evolution of the β-Thymosin/WH2 (WASP-Homology2) Actin-Binding Module 2007).
1. Ce domaine WH2 ou β2/WH2 est trouvé dans un grand nombre de protéines :
- les NPF,
- les protéines de nucléation (Tandem-monomer-binding nucleator) comme Spire et Cordon-Bleu,
- la thymosine-β4.
2. Ce domaine a été trouvé chez les invertébrés, et en particulier dans une protéine appelée Ciboulot, qui joue un rôle majeur dans la croissance axonale au cours du développement cérébral lors de la métamorphose de la drosophile.
- Ciboulot contient 3 répétitions thymosine-β.
- Ciboulot se lie à l'actine comme la thymosine-β4, mais active également la polymérisation de l'actine, par sa première répétition (D1) qui a la même action que la profiline toute entière (Structure, Function, and Evolution of the β-Thymosin/WH2 (WASP-Homology2) Actin-Binding Module 2007).
Structure du domaine WH2
WH2 ne contient que 17 à 27 acides aminés (Actin filament nucleation and elongation factors – structure-function relationships 2009).
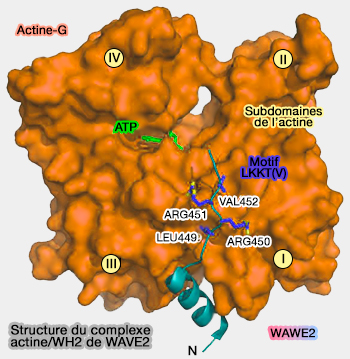
(Figure : vetopsy.fr d'après Dominguez)
1. Sa partie N-terminale forme une hélice-α amphiphilique qui se lie au sillon hydrophobe entre les lobes 1 et 3 de l'actine de l'extrémité barbée (+), comme la plupart les domaines ADF-H, ou G1 et G4 de la gelsoline (The β-Thymosin/WH2 Domain: Structural Basis for the Switch from Inhibition to Promotion of Actin Assembly 2004).
2. Puis, après cette hélice, WH2 présente une région élargie qui monte vers l'extrémité pointée (-) du monomère d'actine.
Cette région, de longueur et de séquence variable ne comprend que 4 résidus hautement conservés, le motif LKKT(V), formé de 4 acides aminés (par exemple, Leucine 449, Arginine 450, Arginine 451, Valine 452 dans la WAVE2).
- L'élément le plus important est la première leucine (L) invariable du motif qui se lie avec l'actine dans la poche hydrophobe (entre les domaines I et III de l'actine - isoleucine 341 et isoleucine 345).
- Puis, en général, on trouve la lysine (K) et l'arginine (R) qui se lient aux résidus acide aspartique 24 et 25, dont le rôle principal est la reconnaissance de l'actine.
- Le quatrième acide aminé (thréonine, valine, alanine ou sérine) s'attache dans une petite poche de l'actine.

L'hélice N-terminale et le motif LKKT(V) constituent le cœur du domaine WH2 (Structural insights into de novo actin polymerization 2010).
Domaine C et A
(Figure : vetopsy.fr)
1. Le domaine C (Cofiline-homology, central ou connecteur) est constitué d'une vingtaine d'acides aminés.
a. Il forme une hélice α amphipathique, i.e. αG, dont la surface hydrophobe active le complexe Arp2/3 pour nucléer les réseaux ramifiés d'actine.
- Il se lie avec la fraction ARPC1/p40 du complexe.
- Comme son nom l'indique, il peut se lier à la cofiline.
b. Son rôle est important dans l'auto-inhibition des WASP car il interagit avec le domaine AI/IS du domaine GBD/PBD ( autoinhibition des WASP).
2. Le domaine A (Acid) se lie aussi à l'Arp2/3 pour son activation et la nucléation de l'actine.
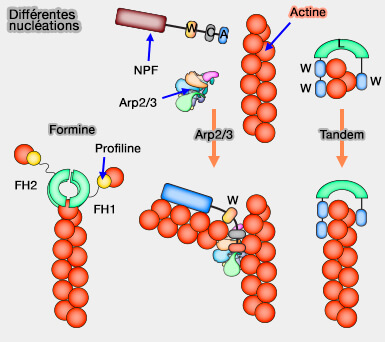
Activation du domaine VCA/WCA
Les protéines de nucléation du filament d'actine catalysent l'étape limitante de la vitesse dans la polymérisation de l'actine, c'est-à-dire la nucléation.
Le complexe ARP2/3 est le seul complexe qui peut générer des réseaux d'actine ramifiés, une architecture qui joue un rôle clé dans des processus mobiles tels que le trafic vésiculaire, l'endocytose/ /exocytose et la motilité cellulaire (The Arp2/3 Regulatory System and Its Deregulation in Cancer 2018).
Le motif WCA induit un changement conformationnel du complexe Arp2/3 et charge une première molécule d'actine sur l'Arp2/3 réarrangé (Regulation of Actin Filament Assembly by Arp2/3 Complex and Formins 2007).
1. Les motifs WCA sont constitutivement actifs, puisqu'ils se replient lors de la liaison de leurs partenaires, l'actine et Arp2/3.
Cette activation a été précisée récemment et nécessite deux sites du complexe Arp2/3 (Cryo-EM structure of NPF-bound human Arp2/3 complex and activation mechanism 2020).
2. Leur régulation spatiale et temporelle dépend de plusieurs processus ( régulations des NPF), afin de réguler l'activation d'Arp2/3, les motifs WCA doivent être masqués
- soit dans une conformation auto-inhibée, comme pour N-WASP,
- soit au sein d'un complexe multiprotéique stable, comme pour WAVE (The Arp2/3 Regulatory System and Its Deregulation in Cancer).
3. Ce domaine permet également l'attachement du réseau d'actine formé à la membrane cellulaire par la suite (Mechanism of Actin Network Attachment to Moving Membranes: Barbed End Capture by N-WASP WH2 Domains 2007).
Domaine riche en proline (PRD ou PRL)
Le domaine riche en proline (PRD ou PRL) est un site pour de nombreuses protéines à domaine SH3 (Src-homology 3) et comprend trois régions distinctes.
1. La première (site de régulation) se lie avec des protéines à :
- domaine SH3, i.e. Abl, Nck, Crk et cortactine,
- domaine WW, i.e. Abl, Src et IRSp53….
2. La deuxième (site de recrutement) comprend trois domaines de liaison à la profiline (GPPPPP ou GP5), et peut donc se lier à plusieurs profilines complexées à l'actine.
3. le troisième (site de chargement) est proche du site de liaison à l'actine-G (GAB) et sert de site de " chargement " de l'actine complexée à la profiline.
Les protéines F-Bar comme Toca-1 (Transactivator Of Cytoskeleton Assembly 1) ou les paralogues CIP4 (Cdc42-interacting protein 4) ou FBP17 (Formin-Binding Protein 17) joueraient un rôle essentiellement dans la déformation membranaire de endocytose clathrine-dépendante (CME), le bourgeonnement des vésicules et la sélection du cargo (The Cdc42-interacting Protein-4 (CIP4) Gene Knock-out Mouse Reveals Delayed and Decreased Endocytosis 2009).
Domaine N-terminal
Les domaines N-terminaux diffèrent selon les familles (The WASP and WAVE family proteins 2009 et The WASP–WAVE protein network: connecting the membrane to the cytoskeleton 2007).
WASP et N-WASP
Les WASP contiennent plusieurs domaines.
Domaine WH1
1. Le domaine WH1 (WASP-homology-1) possède une structure similaire au domaine EVH1 (Ena/VASP homology 1) des Ena/VASP (The WH1 and EVH1 Domains of WASP and Ena/VASP Family Members Bind Distinct Sequence Motifs 2002).
a. Il appartenant à la superfamille des domaines PH, qui peut se lier au PI(4,5)P2 et à de nombreuses protéines par le motif (D/E)FPPPPx(D/E)(D/E).
Ces molécules sont variées, i.e. lamellipodine, Robo dans le guidage axonal, protéines d'adhésions focales, zyxine, vinculine, migfiline et palladine…
(Figure : vetopsy.fr)
b. Ce domaine se lie avec les protéines WIP pour inhiber la dégradation des WASP.
Les mutations dans le syndrome Wiskott-Aldrich sont localisés à ce niveau.
Domaine B
La région B (basique) :
- se lie au PI(4,5)P2,
- entre en synergie avec Cdc42 pour activer les WASP.
Domaine GBD/PBD
Le domaine GBD/PBD est étudié dans un chapitre spécial.
1. Le domaine de liaison aux petites GTPases,, i.e. GBD (GTPase-Binding Domain), appelé aussi PBD (p21 binding domain) peut se diviser en deux domaines.
- a. Le domaine CRIB (Cdc42/Rac interactive binding motif) est la région N-terminale la plus conservée des domaines GBD/PBD.
- b. Le domaine auto-inhibiteur (AI), appelé aussi IS (Inhibitor domain), moins bien conservé et localisé au niveau C-terminal de CRIB, est responsable du maintien des protéines dans un état basal, i.e. auto-inhibé.
Remarque : Le domaine GBD/PBD est quelquefois utilisé comme synonyme de domaine CRIB.
2. Le domaine GBD/PBD auto-inhibe les WASP en étant accolé au domaine VCA.
Il est libéré pour interagir avec les petites GTPases, en particulier de la famille Rho/Rac/CdC42 avec lesquels il interagissent ( accès au domaine VCA).
WAVE
Les WAVE comportent plusieurs domaines (WAVE signalling: from biochemistry to biology 2006).
1. Le domaine N-terminal SHD/WHD (Scar-Homology Domain/WAVE-Homology Domain), est impliqué dans la formation du complexe pentahétéromérique.
- Il se lie d'une part avec Abl et d'autre part avec Brick1, appelé aussi HSPC300.
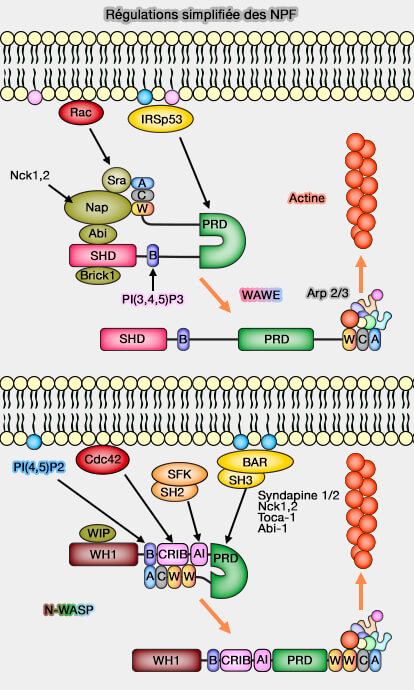
- Abi se lie aussi à Nap, lui-même lié à Sra qui contacte Rac-GTP, ce qui explique l'activation des WAVE par Rac.
2. la région B (basique) se lie au (Activation of the WAVE complex by coincident signals controls actin assembly 2009) :
- PI(4,5)P2 préférentiellement pour WAVE2,
- PI(3,4,5)P3.
Localisation des NPF
WASP et WAVE sont attachés aux phosphoinositides de la membrane cellulaire :
- soit directement en particulier par le domaine B pour WASP etr WAWE,
- soit indirectement à la membrane.
Activant le complexe ARP2/3, elles sont impliquées dans les fonctions liant la membrane et le cytosquelette comme la motilité cellulaire par exemple.
Régulations des NPF
En construction