VAMP7 (également appelé VAMP insensible à la neurotoxine tétanique, TI-VAMP, Tetanus neurotoxin Insensitive-VAMP ou SYBL1, Synaptobrevin-like protein 1), R-SNARE. contenant une arginine (R) dans la couche 0 du motif SNARE, est une v-SNARE (v pour vésiculaire) impliquée dans la fusion vésiculaire, en particulier des lysosomes.
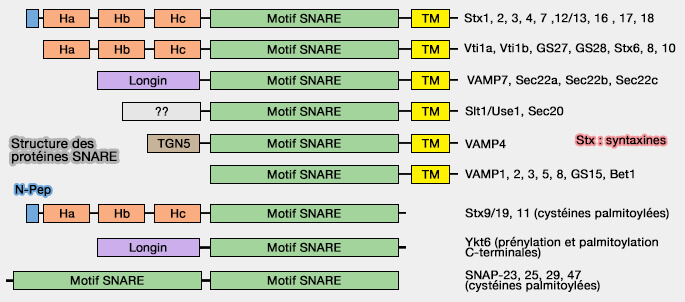
Structure des protéines SNARE
(Figure : vetopsy.fr d'après Hong et Lev)
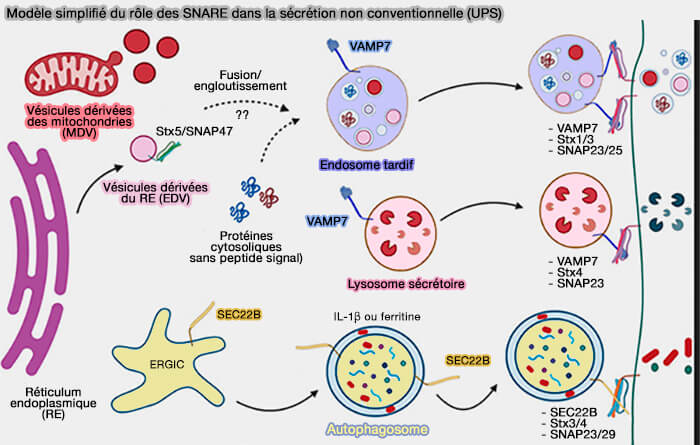
Fusion des lysosomes
1. VAMP7 est impliqué dans la fusion des lysosomes :
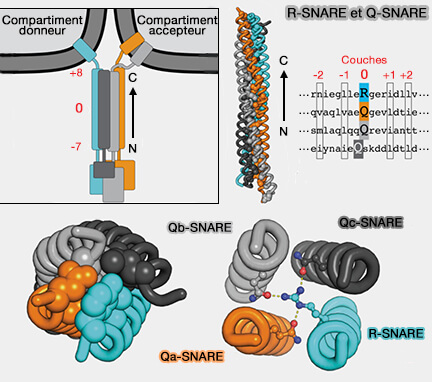
R-SNARE et Q-SNARE
(Figure : vetopsy.fr d'après Baker et coll) avec la membrane plasmique, i.e. accompagné par la syntaxine 4 et SNAP-23,
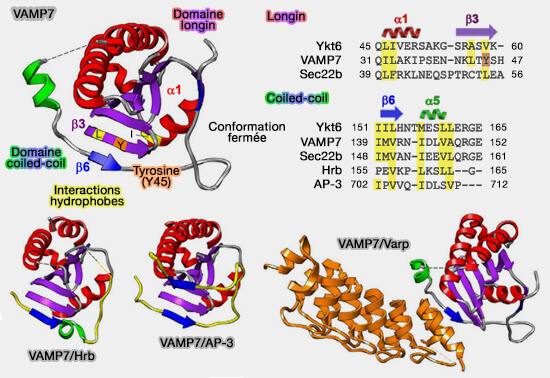
Le domaine longin de VAMP7 est aussi impliqué dans le ciblage de ses sites d'action en se liant à divers partenaires pour pouvoir accomplir ses fonctions.
Ces interactions successives permettent à VAMP7 de circuler de la membrane plasmique vers les endosomes précoces lorsqu'il est lié à Hrb, et de là vers les endosomes tardifs lorsqu'il est lié à AP-3.
VAMP7 et ses interactions
(Figure : vetopsy.fr d'après Daste et coll)
b. Le long de cette route de trafic, VAMP7 doit être conservé sous une forme inactive pour empêcher tout événement de fusion non spécifique selon plusieurs modèles.
VAMP7 et VAMP8 mammaliennes, correspondant à la seule Vamp7 de la drosophile, sont associés à la Qa-SNARE autophagosomale STX1 (Syx17 chez la drosophile) et le Qb/c-SNARESNAP-29 (Ubisnap chez la drosophile) forme le complexe SNARE qui médie directement la fusion autophagosome-lysosome.
Un autre complexe SNARE dans ce processus est composé de YKT6-SNAP-29-STX7.
Toutefois, chez les souris knock-out de VAMP7, on remarque :
Modèle d’interaction entre STX17 et VPS33A de HOPS
(Figure : vetopsy.fr d'après Saleeb et coll)
Le modèle pourrait être le suivant :
a. Avant la fusion, VPS33A du complexe HOPS s'associe à la Ser2 phosphorylée de Stx17.
b. Stx17 est localement déphosphorylé au site de fusion, ce qui modifie la liaison VPS33A et permet la formation du faisceau SNARE.
c. SNAP-29 et VAMP7 peuvent maintenant s'associer à Stx17, formant un complexe trans-SNARE stabilisé par une association VPS33A qui entraîne la fusion membranaire.
d. Après la fusion, le faisceau cis-SNARE persiste jusqu'à ce qu'il soit activement désassemblé.
e. Ser2 de Stx17 est de nouveau phosphorylée pour réengager le domaine 1 de VPS33A avec le peptide N Stx17 pour dissuader d'autres événements de fusion.
Régulation précoce de l'autophagie
VAMP7 est impliqué dans la régulation précoce du processus autophagique comme :
VAMP7 favorise la fusion homotypique des précurseurs ATG16L1 et régule la taille des vésicules ATG16L1, qui sont importantes pour l'expansion de la membrane autophagique.
Le pool ATG16L1-positif de VAMP7 provient également de la membrane plasmique de manière dépendante de l'Hrb/AGFG1, et peut-être également en partie de la voie ATG9A.
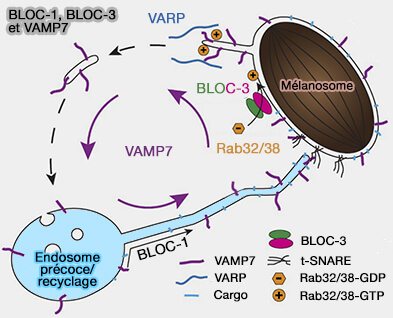
Remarque : les mélanosomes, faisant partie des LRO (Lysosome-Related Organelles), émettent de courts tubules mobiles qui sont enrichis en VAMP7 nécessaire au trafic de fret antérograde dépendant de BLOC-1 ( VAMP7 et complexes BLOC).




 VAMP7 et autophagosomes).
VAMP7 et autophagosomes).