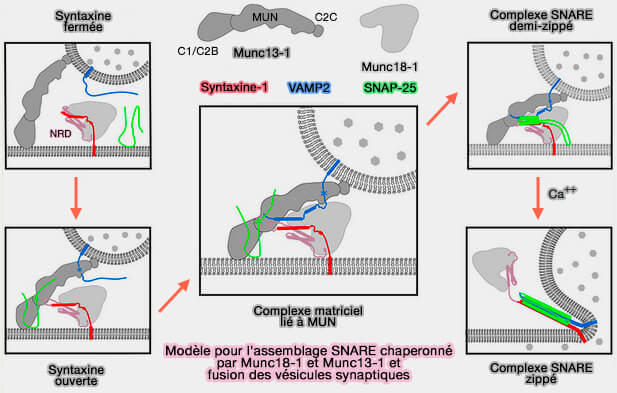
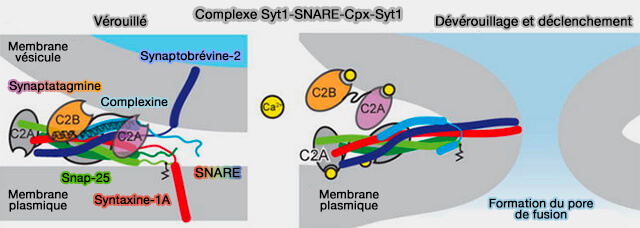
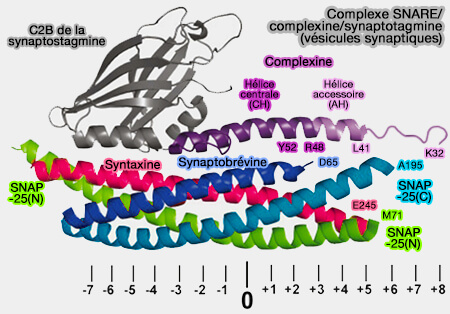
Modèle pour l'assemblage SNARE et fusion des vésicules synaptiques
(Figure : vetopsy.fr d'après Zhang et Hugson)
d. Dans les événements de fusion déclenchés par Ca++, des capteurs comme les synaptotagmines sont déterminants dans la fusion des vésicules synaptiques.
Le RRP (Readily Releasable Pool) est le pool de vésicules qui sont amarrées (dockées), i.e. alignées à moins de 5nm de la membrane présynaptique, et amorcées (primées), i.e. tirées par le complexe SNARE partiellement zippé qui attache la vésicule à la membrane et qui deviennent sensibles au Ca++ et donc, qui sont immédiatement libérées lors d’une stimulation.
Le RP (Reserve Pool), plus grand que le RRP, avec environ trois fois plus de vésicules dans l'hippocampe, représente le pool de vésicules dockées qui doit reformer le RRP quand il a été consommé.
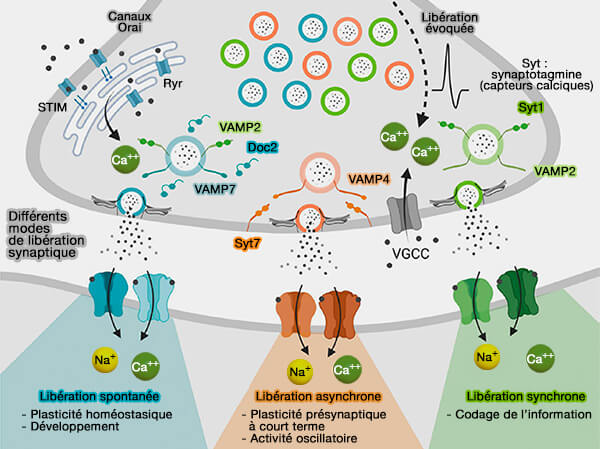
Différents modes de libération présynaptique des neurotransmetteurs
(Figure : vetopsy.fr d'après Guzikowski et Kavalali)
2. Pour ces deux types de vésicules, les protéines SNARE peuvent s'assembler spontanément et provoquer une perte prématurée de vésicules, appelée libération spontanée.
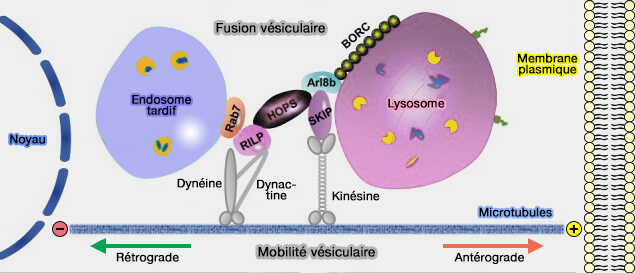
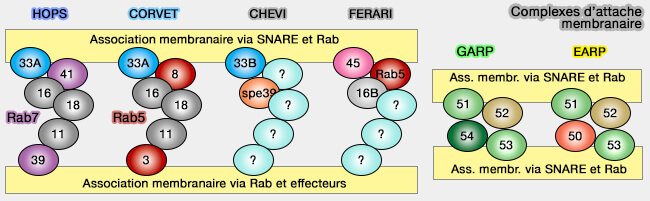
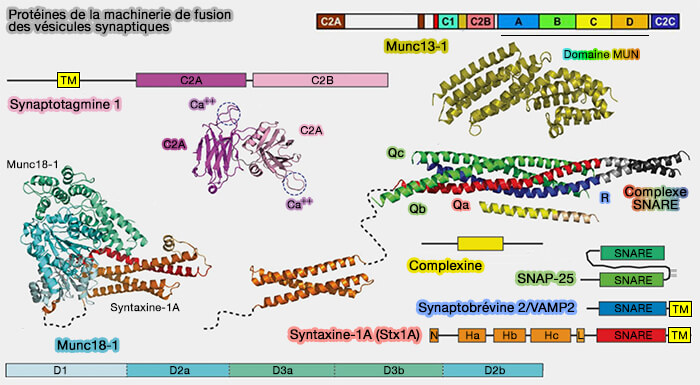
Protéines de la machinerie de fusion des vésicules synaptiques
(Figure : vetopsy.fr d'après Jahn et Fasshauer)
3. Or, la majorité des vésicules ancrées ne doivent pas fusionner avec la membrane plasmique en l'absence d'un bon stimulus, i.e. un mécanisme moléculaire doit exister qui arrête efficacement les vésicules à l'état amarré, permettant un couplage stimulus-sécrétion approprié.
Bien que d'autres mécanismes existent ( régulation du complexe SNARE), les complexines (Cplx) et les synaptotagmines (Syt) fournissent un mécanisme spécifique aux neurones pour la régulation de la sécrétion synaptique.
4. Les mécanismes par lesquels ces protéines régulent l'assemblage et la fusion de SNARE sont encore en cours d'étude, mais plusieurs modèles relient leurs activités biochimiques à des défauts de libération observés chez des mutants perturbant leur fonction.




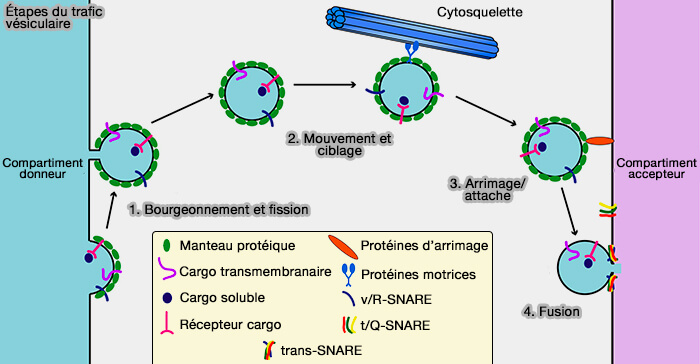
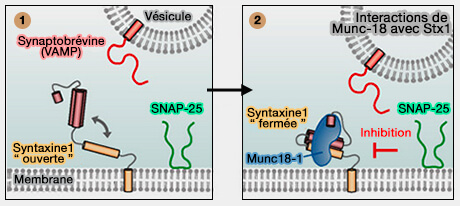

 régulation des SNARE par la phosphorylation de Munc18-1),
régulation des SNARE par la phosphorylation de Munc18-1),