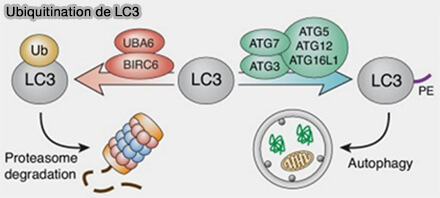
La famille d'Atg8, Atg8 chez la levure, LC3 et GARABAP chez les mammifères, se conjugue de manière transitoire à la membrane autophagosomale par un système de conjugaison de type ubiquitine.
Leur nom vient du fait qu'elles ont été d'abord identifiées comme affectées aux microtubules, puis, par la suite, à l'élongation du phagophore.
2. Trois GABARAP (Gamma-AminoButyric Acid Receptor-Associated Protein), 117 résidus :
GABARAP,
GABARAPL1, 87% d’identité avec GABARAP et 61% avec GATE-16, 30,8% avec LC3A, 29% avec LC3B et 35,8% avec LC3C,
GABARAPL2 ou GATE-16 (Golgi- associated ATPase Enhancer of 16 kDa).
GABARAP a d'abord été trouvé dans les synapses en interaction avec le cytosquelette, puis, par la suite, à la maturation et à la fermeture de l'autophagosome.
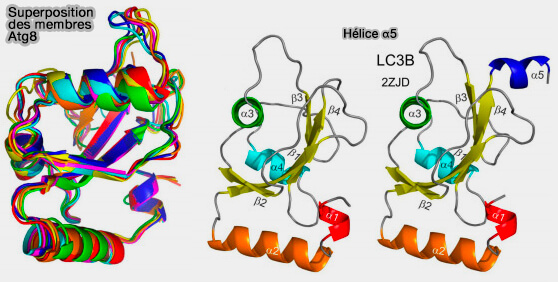
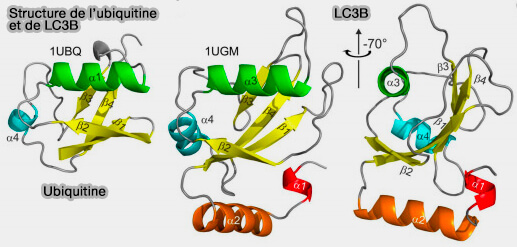
a. Sa structure étroitement repliée, appelée β-grasp fold, est caractérisée par un feuillet-β à 5 brins et une hélice 310 enroulée autour de l'hélice centrale (α1).
b. La protéine précurseur subit une maturation protéolytique pour exposer un résidu de glycine C-terminal, dont le groupe carboxyle peut être conjugué à :
La principale différence structurelle entre les protéines Atg8/LC3/GABARAP et d'autres UBL (Ubiquitin-Like protein) qui détermine également le rôle spécifique d'Atg8/LC3/GABARAP dans l'autophagie, est la présence de deux hélices α supplémentaires situés à l'extrémité N-terminale de l'ubiquitine.
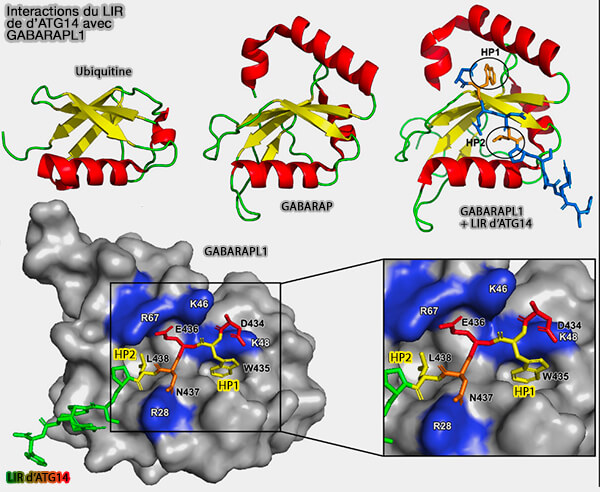
Interactions du LIR d'ATG14 avec GABARAPL1
(Figure : vetopsy.fr d'après Johansen et Lamark)
Par conséquent, cette adaptation structurelle d'Atg8/LC3/GABARAP se reflète dans un ensemble de nouvelles fonctions qui n'ont pas été observées pour d'autres UBL et, par exemple, ces hélices N-terminales sont essentielles pour :
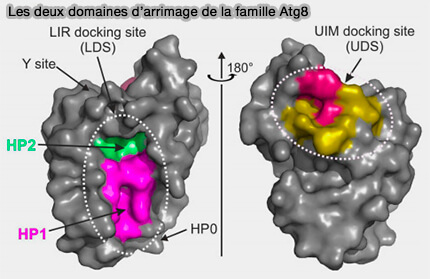
Les deux domaines d’arrimage de la famille Atg8
(Figure : vetopsy.fr d'après Wesch et coll)
2. Deux poches forment le site dit d'arrimage LIR (LDS ou LIR-docking site) et sont impliquées dans une grande majorité d'interactions connues entre les récepteurs d'autophagie sélective (SAR), l'adaptateur et les protéines d'échafaudage avec les Atg8/LC3/GABARAP.
a. Malgré leur flexibilité, les hélices α N-terminales sont spécifiquement alignées sur le noyau de type ubiquitine et forment, avec des résidus du brin β2, une poche hydrophobe profonde 1, i.e. HP1, également appelée site W.
Cette poche se lie préférentiellement à des substances à base d'indole, bien qu'ayant une faible affinité, et accueille généralement de grandes chaînes latérales de résidus aromatiques non polaires au sein des motifs LIR.
b. Une autre poche hydrophobe, i.e. HP2 ou site L, est formée par les résidus exclusivement hydrophobes de l'hélice centrale α3 et du brin β2 (F52, V54, P55, L63, I66 et I67).
Remarque : le LDS occupe la surface opposée à la plaque hydrophobe de l'ubiquitine, i.e. L8-I44-V70.
Les motifs LIR sont étudiés dans un chapitre spécial.
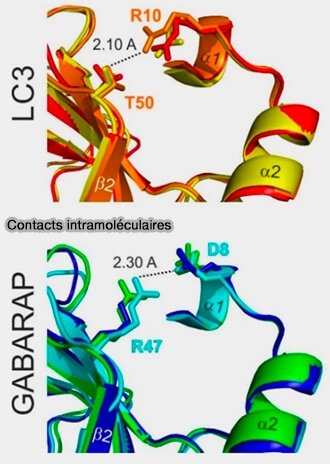
1 La première différence constante entre les sous-types de protéines LC3B et GABARAP est le fait des contacts électrostatiques intramoléculaires pour les résidus 8 et 47 dans Atg8/GABARAP et 10 et 50 dans LC3B, respectivement.
Contacts intramoléculaires
(Figure : vetopsy.fr d'après Wesch et coll)
a. Pour tous les Atg8/GABARAP, la position 8 est occupée par des résidus chargés ou polaires négatifs capables de jouer un rôle d'accepteurs de liaison hydrogène, i.e. Glu, Asp, Gln, Asn, Ser et Thr, tandis que la position 47 est utilisée en permanence pour les résidus électropositifs, i.e. Lys et Arg, servant de donneurs de liaison hydrogène.
b. Pour LC3, il existe des résidus électropositifs, donneurs de liaison hydrogène, en position 10 et des résidus électronégatifs, i.e. accepteurs de liaison hydrogène en position 50.
Ces résidus se rejoignent étroitement et forment des liaisons hydrogène intramoléculaires ou des ponts salins pour stabiliser la structure Atg8/LC3/GABARAP et assurer une orientation appropriée du sous-domaine α N-terminal.
Il est important de noter que ces résidus forment également des contacts intermoléculaires aux résidus dans les motifs LIR et contribuent donc également à la sélectivité des interactions Atg8 avec d'autres protéines.
3. Outre ces deux différences constantes, reflétant la séparation globale des sous-types LC3 et GABARAP, il existe plusieurs points qui régulent la différence dans la reconnaissance des motifs LIR par les membres de la sous-famille.
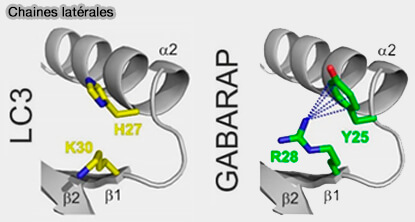
Dans les GABARAP, par exemple, la conformation de Y25 stabilisée par R28 rend les liaisons hydrogène intermoléculaires plus favorables pour l'affinité du GABARAP aux motifs LIR.
Dans le LC3, il y a H27/H27/F34 (LC3A/LC3C) à la position Y25, et par conséquent, les liaisons hydrogène ne pouvaient pas être formées de la même manière.
Modifications post-traductionnelles
En outre, les modifications post-traductionnelles des protéines Atg8/LC3/GABARAPs peuvent affecter l'affinité et la sélectivité des interactions LIR:Atg8/LC3/GABARAP.
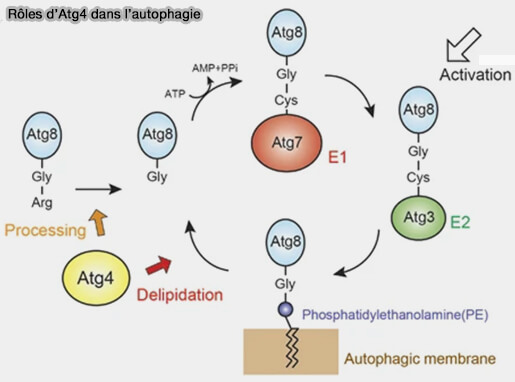
Par exemple, la phosphorylation par TBK1 de LC3C et de GABARAPL2 régule leur délipidation en perturbant spécifiquement la capacité des protéines ATG4 à reconnaître ces substrats ( TBK1 et Atg4).




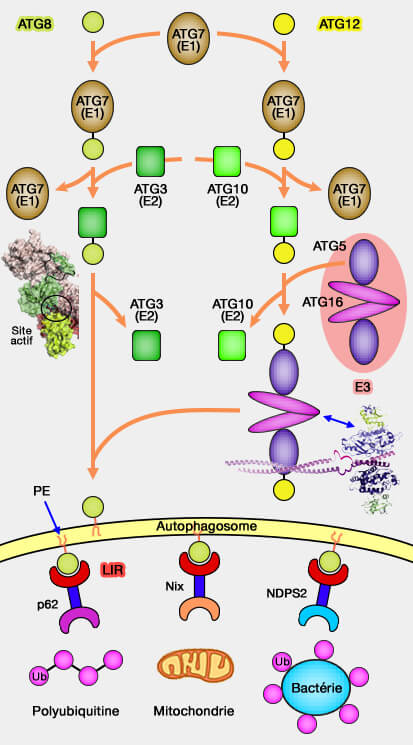


 les deux systèmes de type ubiquitine) :
les deux systèmes de type ubiquitine) :