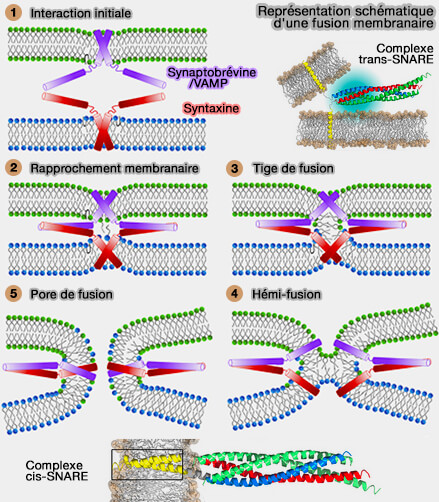
Les fusions membranaires dans le trafic vésiculaire, i.e. voie sécrétoire et endocytose, doivent être à la fois spatialement et temporellement réglementées, afin que le cargo puisse être fidèlement livré au bon organite.
1. Les changements topologiques membranaires se produisent par des événements de fusion qui font intervenir :
Ces transporteurs (carriers) peuvent être ronds ou tubulaires et, quelle que soit leur morphologie, ils doivent contenir ou acquérir la machinerie nécessaire pour fusionner avec l'organite accepteur.
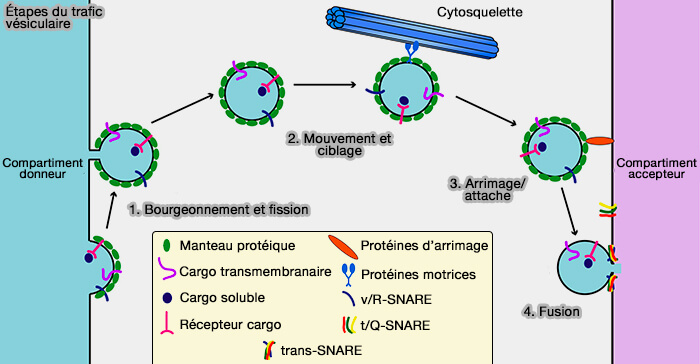
Étapes du trafic vésiculaire
(Figure : vetopsy.fr d'après Cai et coll)
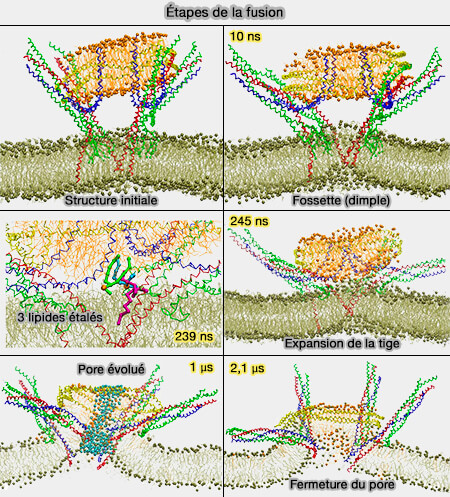
2. La fusion membranaire est tout sauf un processus spontané.
Les bicouches lipidiques intracellulaires sont constituées de lipides membranaires qui portent des groupes de tête hydrophiles et des queues hydrophobes.
Elles forment ainsi un noyau hydrophobe avec de l'eau sur la surface luminale ou cytoplasmique. Tout facteur de fusion doit donc contourner cet obstacle.
Les barrières énergétiques pour la fusion des vésicules comprennent la déshydratation des groupes de tête des phospholipides (PL) et l'encombrement stérique, i.e. pour que deux membranes opposées entrent en contact, les protéines doivent être écartées.
La répulsion électrostatique entre la vésicule et la membrane plasmique bloquerait l'amarrage et la fusion des vésicules et contribuerait à cette barrière énergétique.
Machinerie de fusion
La fusion des vésicules nécessite une machinerie de fusion conservée par l'évolution et composée de plusieurs protéines.
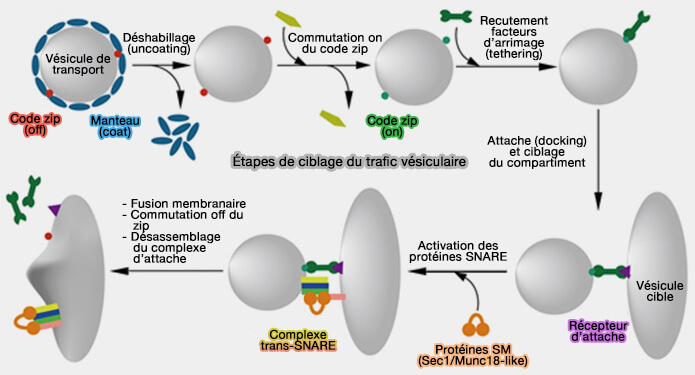
Étapes de ciblage du trafic vésiculaire
(Figure : vetopsy.fr d'après Koike et Reinhard)
Le trafic vésiculaire est incroyablement complexe et les organites sont entourés de milliers de vésicules qui partent et viennent constamment.
Les cellules doivent donc savoir d'où provient une vésicule donnée, i.e. du réseau trans-Golgi (TGN) ou de la membrane plasmique par exemple, et où elle doit être délivrée, i.e. à l'endosome tardif ou à la membrane plasmique par exemple.
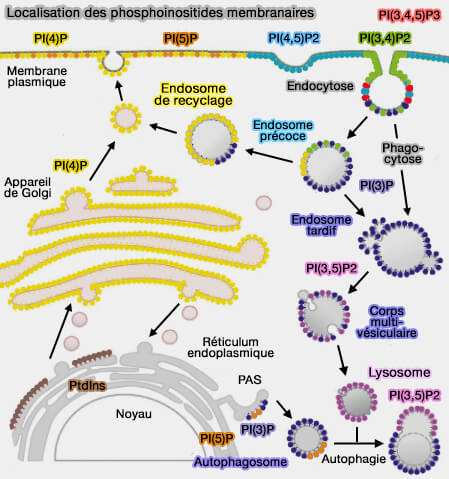
Localisation des phosphoinositides membranaires
(Figure : vetopsy.fr d'après Cernikova et coll)
Pour atteindre leur destination intracellulaire, les vésicules de transport doivent comporter des signaux de ciblage qui sont reconnus par des facteurs de liaison dans le cytoplasme et qui relient les vésicules à leur compartiment de destination respectif.
Composants de la machinerie de fusion
Les fusions spécifiques sont le plus souvent traitées dans des chapitres particuliers aux organites.
des protéines dynamiques qui font la navette entre un état cytosolique inactif, Rab liée au GTP (Rab-GTP), et un état actif attaché à la membrane, i.e. Rab liée au GDP (Rab-GDP), en interaction avec leurs régulateurs GEF, GAP et GDI.
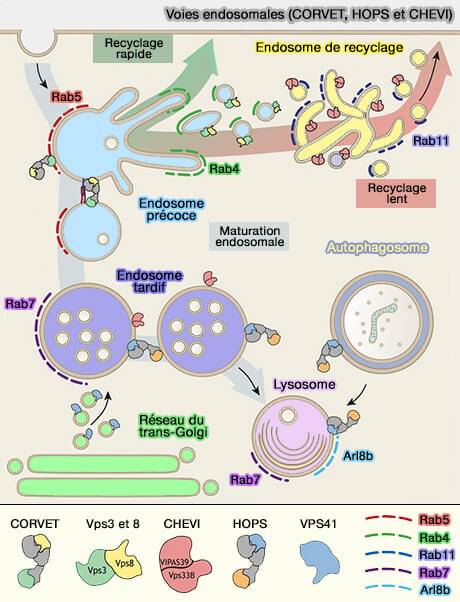
Voies endosomales (CORVET, HOPS et CHEVI)
(Figure : vetopsy.fr d'après van der Beck et coll)







 problèmes de sémantique) :
problèmes de sémantique) :